
Remember me


ADHD is a complex disorder that involves the interaction of many different genes and proteins, resulting in specific behavioral characteristics. Studies have shown that manipulating a single gene or protein can impact other genes and proteins, leading to symptoms such as inattention, hyperactivity, and impulsivity. Our previous study suggests that the predominantly inattentive THRSP-OE mice have intrinsic impairments in the Wnt signaling pathway, as reported through hippocampal proteomics [7], which predisposes this animal model to inattention.
To extend our findings, we conducted a proteomic study using the striatum in mice. The striatum is an important brain region for attention and is affected in individuals with ADHD [18]. It regulates the release of dopamine, a neurotransmitter that plays a vital role in attention, motivation, and reward. Interestingly, people with ADHD have lower levels of dopamine in the striatum, a dysfunction called “hypo-dopaminergic trait,” which may contribute to their attentional deficits [19]. Moreover, the striatum is connected to other brain regions involved in attentional processes, such as the prefrontal cortex [20]. Indeed, dysfunctions in the striatum and its connections to other brain regions can also contribute to attentional deficits in people with ADHD.
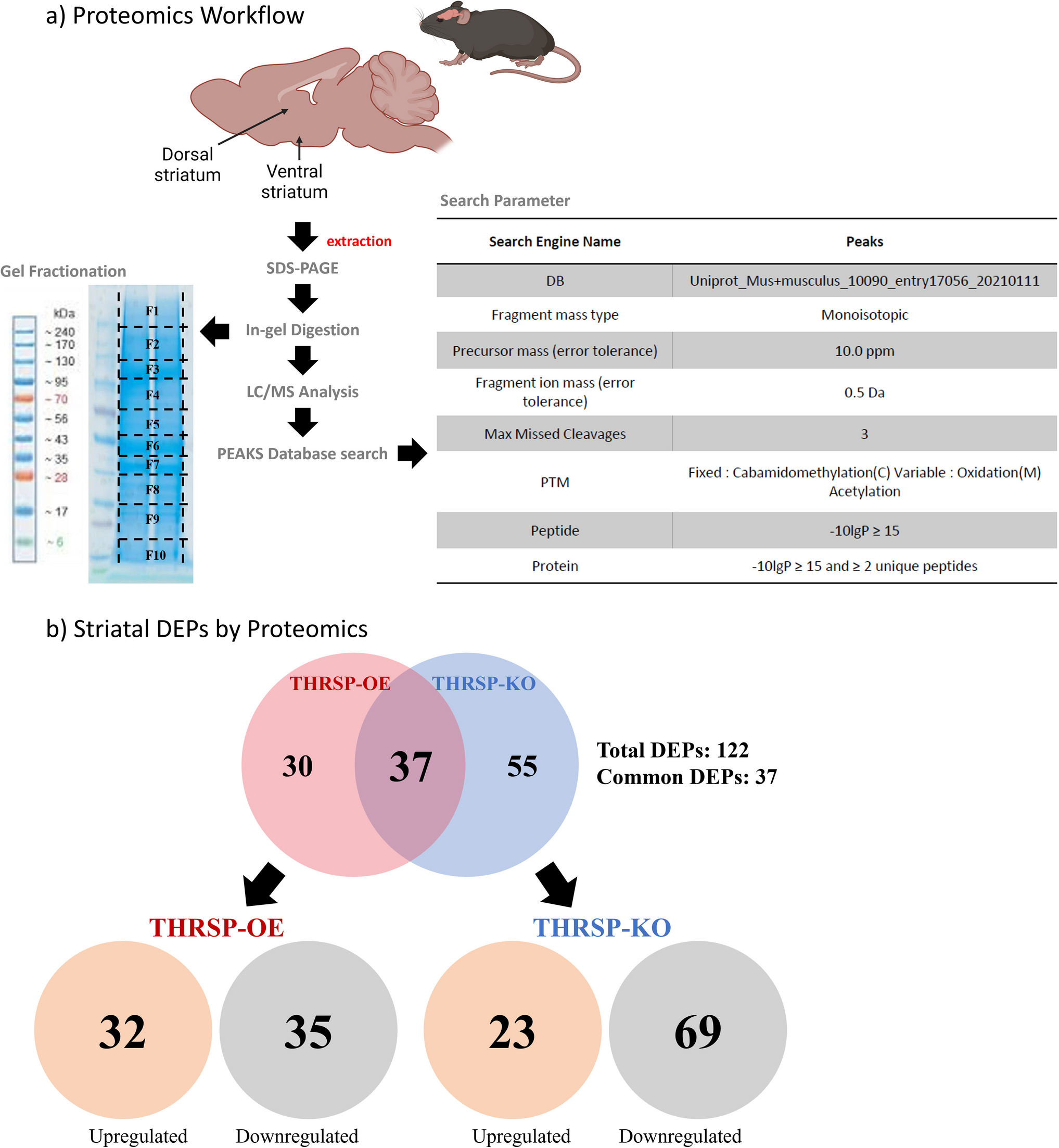
In the present study, a total of 122 DEPs were identified in THRSP-OE and THRSP-KO relative to WT mice (Fig. 1b; Table 1) and showing 37 common DEPs. Out of these, 32 and 35 proteins were upregulated and downregulated in THRSP-OE, respectively, whereas 23 and 69 proteins were upregulated and downregulated in THRSP-KO, respectively. These findings suggest that the overexpression or knockout of the THRSP gene can have a significant intrinsic impact on the expression of other proteins in mice. Using this information, we applied the PANTHER classification system to identify the GO biological processes involved in the DEPs. GO analyses of upregulated (Fig. 2a) and downregulated (Fig. 2b) proteins in THRSP-OE mice identified variations in vesicle-mediated transport, cellular localization, cellular processes, and transport, among others. On the other hand, the GO analyses of upregulated (Fig. 3a) and downregulated (Fig. 3b) proteins in THRSP-KO mice identified involvement in the regulation of vesicle-mediated transport, localization, and biological processes, among others.
These cellular and biological processes show the possibility of impaired synaptic signaling involvement, particularly with THRSP-OE mice, which could be related to the established predominant inattention observed in this ADHD mouse model [9]. However, it must be examined whether synaptic signaling is impaired in response to its transgenic nature. Therefore, using the DEPs both in the THRSP-OE and THRSP-KO mice, we conducted enrichment analysis using PANTHER pathways and identified enriched pathways, including synaptic vesicle trafficking and receptor-mediated signaling pathways (Table 2), including dopamine receptor-mediated signaling pathways known to be involved in ADHD [21]. Moreover, it is interesting to observe that the thyrotropin-releasing hormone receptor signaling pathway was already identified, which further supports the role of thyroid hormone signaling in ADHD [22, 23], which we previously evaluated in the same mouse model [8].
Using the enriched pathways identified, we have further scrutinized what proteins were involved in the presented cellular and biological pathways, and we were intrigued to find out that most recurring sets of proteins involved were those with contributory roles in plasma membrane and synaptic vesicle regulation (Table 3), particularly the Snap25 protein, a member of the family of proteins that make up the soluble N-ethylmaleimide-sensitive factor attachment protein receptors (SNARE) complex, known to have direct links to neurotransmitter regulation including dopamine [24] and previously identified to be involved in ADHD pathology both in humans [2, 25, 26] and animal models [27,28,29].
It has been shown that the DEPs in both the THRSP-OE and THRSP-KO mice belong to the SNARE proteins, member traffic proteins, and G-proteins classification (as shown in Figs. 2c, 2 d, 3c, and 3 d). This indicates that the changes in the THRSP gene, especially in THRSP-OE mice, have intrinsic effects on different molecular functions and pathways, particularly transduction and signaling (as outlined in Tables 4 and 5), among others, which may be responsible for the predominant inattention in this ADHD model.
Furthermore, the total DEPs were also analyzed using STRING, showing protein–protein interaction networks restricted to high-confidence (0.9) interaction thresholds only. The protein networks from THRSP-OE and THRSP-KO mice revealed the high interaction of Snap25 (Fig. 4) among other SNARE complex-related proteins. These critical findings provide evidence of differences between the two transgenic mice, particularly how Snap25 is among the five differentially regulated proteins (Table 5) in THRSP-OE and THRSP-KO mice to which the THRSP-OE mice show upregulation of Snap25. In contrast, THRSP-KO mice show downregulation of this protein. Of the DEPs identified in the study, the Snap25 protein is of particular interest given its important function in the SNARE protein complex by maintaining the complex’s assembly and stability and proper synaptic transmission and brain function, if impaired, can lead to impaired neurotransmitter release (including dopamine) and eventually inducing neuropsychiatric disorders [30], such as ADHD. Overall, the current focus is on understanding how the upregulation of Snap25 affects the regulation of the SNARE protein complex and its targets and evaluating its contributory effects on the established ADHD-PI-like behaviors observed in THRSP OE mice.
The Differentially Regulated Proteins in THRSP OE MiceNotably, five proteins were identified to be differentially regulated (Table 6) among all DEPs, meaning that these protein levels were opposing between the two mouse groups (THRSP-OE vs THRSP-KO). For instance, we observed that Dennd4c, Snap25, and Vwf proteins were upregulated in THRSP-OE mice, while these were downregulated in THRSP-KO mice. Conversely, Dpysl3 and Gnai3 proteins were downregulated in THRSP-OE and upregulated in THRSP-KO mice. Given these findings, we analyzed the differentially regulated proteins identified in striatal proteomics with a particular focus on the THRSP-OE mice, given that this animal model has an established ADHD-PI-like behavior and is the target of the study. Therefore, at this stage, we have conducted the mRNA analysis via RT-qPCR in dennd4c, snap25, dpysl3, vwf, and gnai3 genes only in the ADHD-PI model, THRSP-OE mice.
It is worth noting that all five differentially regulated proteins play crucial roles in nervous system function, including neurotransmission, neuronal connectivity, and blood flow regulation. Dendd4c, also known as Ras-related protein Rab-10 (Rab10), is a protein that regulates endocytosis [31], a process that helps control the number of receptors on the surface of cells, including neurons and also found to regulate dendritic branching by balancing dendritic transport [32]. Snap25 is a SNARE protein family member that plays an essential role in neurotransmitter release at the synapse [5], a process necessary for communication between neurons. Dpysl3, also known as collapsin response mediator protein 4 (Crmp4), is involved in axon guidance [33], which helps ensure proper neuronal connectivity during development. Vwf, or von Willebrand factor, is involved in blood clotting, found to promote blood–brain barrier flexibility [34]. Finally, Gnai3 is a G protein subunit (inhibitory alpha subunit) that is a component of the G protein complex (alpha, beta, and gamma subunits), which plays a crucial role in signaling pathways regulating neurotransmitter release and synaptic plasticity. As a key regulator of G protein-coupled receptor signaling, Gnai3 has been implicated in the modulation of neuronal excitability and synaptic transmission, and its dysregulation has been linked to various neurological disorders [35].
Results show increased levels of dennd4c and snap25 genes in the striatum of THRSP-OE mice compared to the WT mice (Fig. 5a, b). Moreover, there was a decreased expression of the dpysl3 gene (Fig. 5c) in the striatum of THRSP-OE mice compared to the WT mice. This shows similar trends with the expression levels of the Dendd4c, Snap25, and Dpysl3 protein levels identified in the striatal proteomics. However, while we found similarities in the expression levels of protein and genes for the three mentioned above, we failed to see significant changes in the expression levels of the vwf and gnai3 genes (Fig. 5d, e) in the striatum of THRSP-OE mice relative to the WT mice. Nonetheless, we found sustained expression levels of the snap25 gene which is among the most important findings in this study, given its previous involvement in ADHD pathology [4, 26, 27, 29].
The SNARE ComplexOur study has uncovered novel and sustained changes in the snap25 gene expression levels in THRSP-OE mice. This discovery prompted us to delve deeper and examine the responses of the other genes involved in the SNARE complex and voltage-dependent Ca2 + channels (VDCC). The SNARE complex and VDCC, both of which are crucial in neurotransmitter release, are formed by the interaction of several proteins, including Vamp2 (located on the vesicle membrane), Snap25, and Stx1a and Stx1b (located on the presynaptic membrane) [36]. Functionally, this complex is responsible for the docking of synaptic vesicles at the presynaptic membrane and the subsequent fusion of the vesicle membrane with the presynaptic membrane through the recruitment of necessary proteins, such as Syt1, Munc13, and Munc18, leading to neurotransmitter release into the synaptic cleft. VDCC, on the other hand, allows Ca2 + influx into the presynaptic terminal, which triggers the release of neurotransmitters by the SNARE complex.
Given the innate upregulation of the Snap25 protein identified in the proteomics analysis and the resultant snap25 gene overexpression by RT-qPCR, one would assume that the SNARE complex, where this gene or protein is highly involved, may be altered. However, it is interesting that the key players in the SNARE complex, namely the stx1a, stx1b, and vamp2 genes, were not significantly altered (Fig. 6a, b, c). However, we only found a slightly increased expression level of the stx1a gene but a slightly reduced stx1b gene expression level relative to the wild-type. Moreover, we found no slight changes in the expression level of the vamp2 gene. This is a quite intriguing result given that both the Stx1b and Vamp2 proteins were among the DEPs identified in the proteomics in which both the Stx1b (STN ratio = 1.5573; p = 0.0023) and Vamp2 (STN ratio = 2.0911; p = 0.0004) proteins were upregulated relative to the wild-type (refer to Table 1). The observed discrepancy between the protein and gene expression data, particularly between Stx1b protein and stx1b gene and Vamp2 protein and vamp2 gene, suggests that post-transcriptional or post-translational modifications may be involved in regulating the expression or function of these proteins. Also, we observed significant reductions in syt1 and munc13 genes, but the munc18 gene slightly increased, though it did not reach a significance level (Fig. 6d, e, f). The significant reductions in syt1 and munc13 gene expression suggest that changes in snap25 expression may have downstream effects on other genes involved in the SNARE complex. These reductions in syt1 and munc13 gene expression could potentially lead to changes in the SNARE complex’s activity and the neurotransmitter release process. However, it is important to note that the downstream effects of changes in snap25 gene expression on the function of the SNARE complex and neurotransmitter release are complex and may involve multiple genes and pathways.
Furthermore, two VDCC-related genes, namely cacnb2 and cacnb3 genes, were overexpressed in THRSP-OE mice (Fig. 6g, h). The exact mechanism by which Snap25 regulates the expression levels of cacnb2 and cacnb3 genes has yet to be fully understood. However, it is thought to be since Snap25 plays a critical role in regulating VDCC and calcium-dependent neurotransmitter release. The increase in the expression levels of cacnb2 and cacnb3 genes, when snap25 is overexpressed, may be a compensatory mechanism to enhance the activity of the channels and promote efficient neurotransmitter release [37].
These findings underscore the complexities of the regulatory mechanisms involved in SNARE complex formation and function and the need for further studies to fully understand the molecular mechanisms underlying the observed effects of snap25 gene overexpression in THRSP-OE mice on the SNARE complex and neurotransmitter release, particularly the dopamine. The unexpected results and the potential involvement of post-transcriptional or post-translational modifications in regulating the expression or function of these proteins further emphasize the intricacy of these processes.
Dopamine Neurotransmission in THRSP-OE MiceDopamine neurotransmission plays a critical role in Snap25 function because dopamine is a key neurotransmitter in several important physiological processes, including reward, motivation, and movement. Dopamine is released into synapse by presynaptic neurons. It binds to specific receptors postsynaptic, activating downstream signaling pathways that regulate various cellular activities. The SNARE complex regulates dopamine release, which includes Snap25 as a key component [5, 35]. Snap25 plays a crucial role in the synaptic vesicular docking and fusion with the presynaptic membrane, allowing dopamine release into the synapse. Therefore, changes in Snap25 expression levels can have downstream effects on dopamine neurotransmission, potentially leading to alterations in behavior. Indeed, we found a reduced dopamine D1 receptor (D1R) binding (Fig. 7a) in THRSP-OE mice with no significant change in dopamine D2/D3 receptor (D2/3R) binding (Fig. 7b) relative to wild-type mice. Interestingly, this reduction in D1R binding was accompanied by low dopamine levels (Fig. 7c), which was rescued by methylphenidate (5 mg/kg) administration for seven days.
Fig. 7
The dopamine receptor mediated signaling in THRSP-OE mice. Binding affinity towards dopamine D1 receptors and D2/3 receptors were analyzed in striatal samples of THRSP-OE mice (n = 5 samples/group; a unpaired t-test: t = 13.72, df = 4, p < 0.001; b unpaired t-test: t = 0.8739, df = 4, p = 0.4315). Moreover, dopamine levels in the striatum of mice treated with MPH (5 mg/kg) for 7 days were also analyzed (n = 8 samples/group; c one-way ANOVA, F (3, 28) = 10.6, p < 0.001). Values are presented as the mean ± standard error of the mean (SEM). *p < 0.05 ***p < 0.001, by unpaired t-test relative to WT and one-way ANOVA with Tukey’s multiple comparisons relative to vehicle (VEH)-treated WT and THRSP-OE mice
These findings show that the reduced D1R binding and low total striatal tissue dopamine concentrations in the striatum of the THRSP-OE may be a possible result of the altered function of the SNARE complex, leading to reduced dopamine release and signaling in the brain of THRSP-OE mice, which supports the ADHD-PI-like behaviors observed in this animal model. Additionally, a reduction in tyrosine 3-monooxygenase (commonly known as tyrosine hydroxylase), the rate-limiting enzyme in dopamine biosynthesis, may contribute to decreased dopamine levels by restricting its initial synthesis from L-tyrosine, further exacerbating the effects of impaired vesicular release. Notably, both THRSP-OE and THRSP-KO mice exhibit reduced expression of tyrosine 3-monooxygenase (see Table 1; DEP no. 36), with THRSP-OE mice showing a −2.2049-fold change (STN ratio), and THRSP-KO mice displaying an even more pronounced reduction (−4.2800 STN ratio). These findings suggest that THRSP dysregulation impairs dopamine synthesis regardless of whether THRSP is overexpressed or knocked out. However, it is important to note that the dopaminergic system in the brain is complex and involves multiple signaling pathways and receptors. In the case of ADHD, it has been proposed that there may be a dysfunction in the cortico-striatal networks, leading to the characteristic symptoms of the disorder [38,39,40]. Nonetheless, we have once again identified that THRSP-OE mice, an animal model for ADHD-PI, have impaired dopamine neurotransmission, as evidenced by low D1R binding and dopamine concentrations. Taken together, these results reinforce the relevance of THRSP-OE mice as a robust and mechanistically informative model for investigating the dopaminergic underpinnings of ADHDPI. However, continued investigation is warranted to fully elucidate how THRSP dysregulation alters dopamine-related molecular pathways and broader network-level processes underlying ADHD pathophysiology.
EEG Changes in THRSP-OE MiceRecent studies have used EEG to measure possible changes in brain wave activity to understand better the neural mechanisms underlying ADHD [41]. In line with this approach, the current study aimed to investigate EEG changes in early adult THRSP-OE mice, an animal model of ADHD-PI. Our previous finding identified EEG changes in the same strain at four weeks, which can be considered childhood age [8]. Therefore, the primary objective of this study is to report the EEG changes in THRSP-OE mice at an early adult age, which can facilitate a better understanding of the neural mechanisms underlying ADHD pathology between two crucial stages, childhood, and adulthood.
The study revealed significant findings: THRSP-OE mice exhibit innate high theta and beta waves (Fig. 8a, c) compared to the wild-type control. Acute methylphenidate administration was found to reduce the theta (Fig. 8b) but not beta waves (Fig. 8d) in mice. Interestingly, there were no changes in the theta and beta waves between THRSP-OE and wild-type mice during the 7 th day recording (Fig. 8e, g). Equally intriguing, there were no changes in theta or beta waves in the THRSP-OE mice between acute and repeated (7 days) dose of 5 mg/kg of methylphenidate administrations (Fig. 8f, h), although a slightly reduced theta and beta waves were observed, respectively, but failed to reach significance level. It is worth noting that our EEG study involved administering varying doses of methylphenidate (2, 5, and 10 mg/kg), but we primarily focused on the 5 mg/kg dose, as it was the only dose that yielded a significant difference between WT and THRSP-OE mice in our analysis. This finding is consistent with our previous research [6], which demonstrated that MPH administration at a dose of 5 mg/kg improves inattention and striatal dopamine-related gene (i.e., dopamine transporter, tyrosine hydroxylase, D1R, and D2R) expression in THRSP-OE mice. However, 7 days of methylphenidate administration increased the EEG peaks in THRSP-OE mice (Fig. 8l), which is an important finding. A representative frequency band was also shown (Fig. 8m). The observation of increased EEG peaks in THRSP-OE mice following 7 days of methylphenidate administration suggests that 5 mg/kg of methylphenidate may have a positive effect on the neural activity in THRSP-OE mice. This result is consistent with the known effects of methylphenidate on increasing neural activity and improving cognitive function in individuals with ADHD [42, 43]. Moreover, the theta and beta waves during the 1st and 7th day following methylphenidate administration (Fig. 8i, j) did not induce significant changes. However, the observation of reduced TBR is a significant finding in this ADHD-PI mouse model following 7 days of methylphenidate administration (Fig. 8k). Further, findings on the effects of methylphenidate in WT mice can also be found in Supplementary Fig. 1.
Fig. 8
The EEG measurements (theta, beta) in THRSP-OE mice exposed to acute and repeated MPH (5 mg/kg). Mice (n = 6 samples/group) were monitored in an EEG apparatus after single (1 day) VEH and MPH administrations a Theta ((WT (VEH) × THRSP-OE (VEH); unpaired t-test, t = 3.23, df = 5, p = 0.023); b Theta ((THRSP (VEH) × THRSP-OE (MPH); unpaired t-test, t = 5.05, df = 5, p = 0.004); c Beta ((WT (VEH) × THRSP-OE (VEH); unpaired t-test, t = 2.76, df = 5, p = 0.040); d Beta ((THRSP (VEH) × THRSP-OE (VEH); unpaired t-test, t = 2.22, df = 5, p = 0.077), the repeated (7 days) VEH and MPH administrations; e Theta ((WT (VEH) × THRSP-OE (VEH); unpaired t-test, t = 109, df = 5, p = 0.918); f Theta ((THRSP (VEH) × THRSP-OE (MPH); unpaired t-test, t = 0.645, df = 5, p = 0.547); g Beta ((WT (VEH) × THRSP-OE (VEH); unpaired t-test, t = 0.0652, df = 5, p = 0.951); h Beta ((THRSP (VEH) × THRSP-OE (VEH); unpaired t-test, t = 0.767, df = 5, p = 0.478), and the comparison between 1 and 7 days of MPH effects in the EEG of THRSP-OE mice i Theta (THRSP-OE (MPH) (1 day × 7 days); paired t-test, t = 1.38, df = 5, p = 0.227); j Beta (THRSP-OE (MPH) (1 day × 7 days); paired t-test, t = 1.28, df = 5, p = 0.256); k Theta/Beta ratio (THRSP-OE (MPH) (1 day × 7 days); paired t-test, t = 2.72, df = 5, p = 0.042). l EEG peaks were also analyzed in THRSP-OE MPH-treated mice (1 day × 7 days); paired t-test, t = 3.71, df = 5, p = 0.014) and m graphical representations of the EEG amplitudes are also presented. Values are presented as the mean ± standard error of the mean (SEM). *p < 0.05 and **p < 0.01, by unpaired t-test relative to treatment and paired t-test for day effects
The TBR, a reliable measure of ADHD, is diagnosed based on the presence of a higher TBR in individuals with ADHD as compared to those without the disorder [17]. This ratio also resembles the pre-methylphenidate TBR in THRSP-OE mice (Fig. 8k), indicating the potential for improvements in TBR in this ADHD-PI animal model following 7 days of methylphenidate administration, a first-line drug for ADHD, a result supported by prior studies on the improvement of EEG spectral powers in children and adults with ADHD [44, 45]. In 2013, the US Food and Drug Administration (FDA) approved the use of EEG-based methods to diagnose ADHD, such as the Neuropsychiatric EEG-Based Assessment Aid (NEBA) System [46], a noninvasive neurophysiological scan test that gauges an increased brain oscillatory TBR in ADHD. These implications underscore the potential of our findings in advancing the understanding and treatment of ADHD.
Furthermore, our findings suggest that the THRSP-OE mouse model may be a useful tool for investigating the neural mechanisms underlying ADHD and the effects of methylphenidate administration and could potentially be used to inform the development of more effective treatments for ADHD in humans. The theta waves are associated with daydreaming and inattention, while beta waves are associated with focus and attention. Therefore, a higher ratio of theta to beta waves indicates greater inattention (and distractibility), a hallmark symptom of ADHD, and the specific symptom established and modeled by the THRSP-OE mice, an ADHD-PI animal model.
Comments (0)