
Remember me

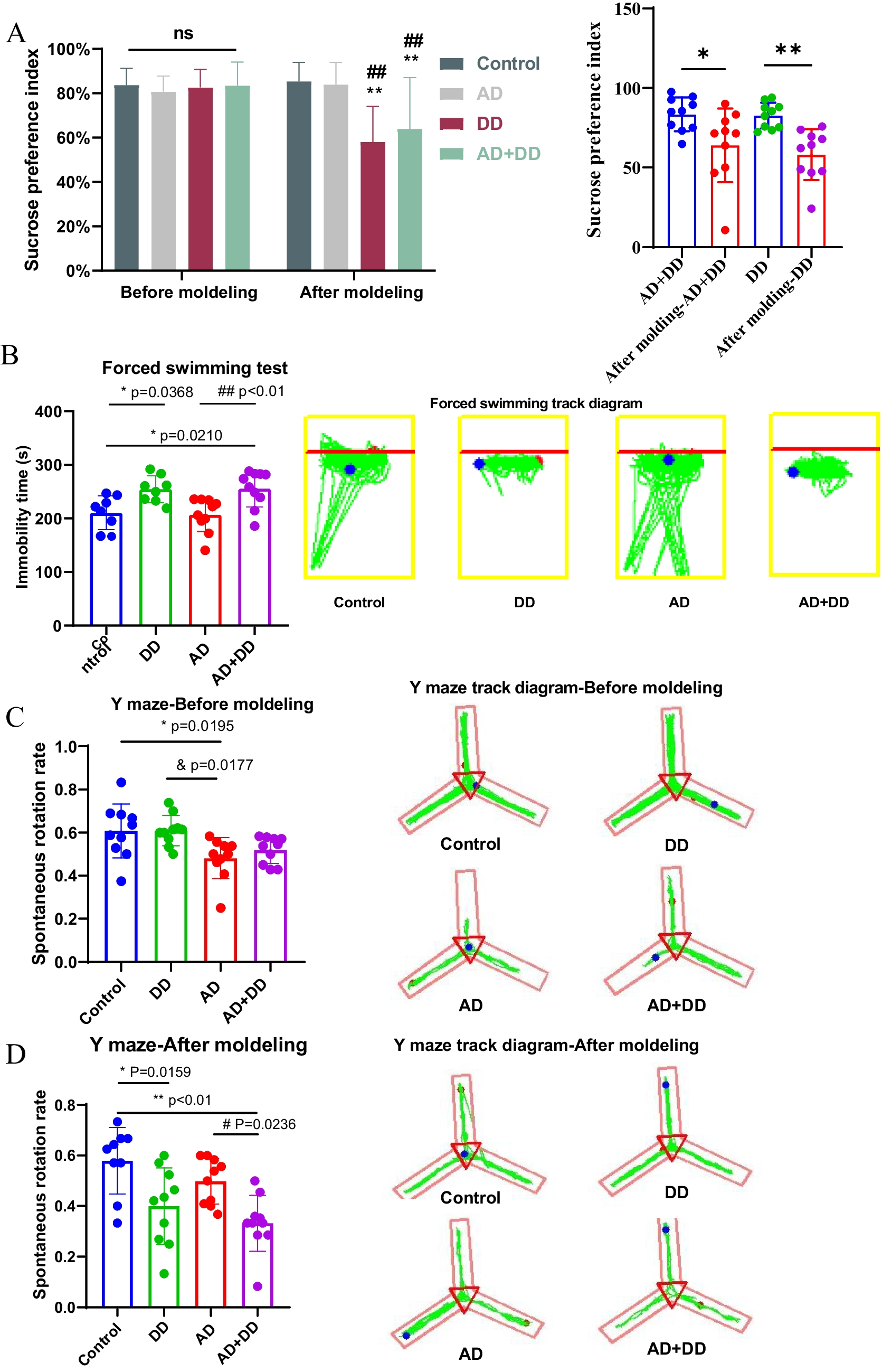

Out of 260 admissions, 258 patients were initially considered for the study. However, 136 were excluded based on inclusion criteria. Nineteen patients opted out. In all, 77 patients consented to participate, among which 36 patients did not experience seizures and three withdrew. Four samples were removed from analysis as events were determined to be non-epileptic, and one additional sample was removed as the seizure type remained unclear after vEEG analysis. Six samples failed for quality check (Suppl. Figure 1). A total of 27 sets of patient samples with either focal or generalized seizure as determined by EEG and chart review by a board-certified epileptologist were included in this study (Table 1).
Table 1 Patient data included in the studyDifferential Expression Profiles Following Seizure Type Paired with BaselineWe identified 50,081 novel transcripts in our dataset that aligned with the GRCh38 human genome, with 67.7% mapping to novel genes and 32.3% associated with known genes (Fig. 1a). Among these, 3191 novel isoforms derived from novel genes showed no overlap with the GRCh38 reference genome (Fig. 1b). We then analyzed differential transcript expression following generalized and focal seizures, comparing post-seizure samples to their respective baseline samples (Table 1). In patients with focal seizures, we identified 41 differentially expressed transcripts between baseline and 4–6 h post-seizure samples (26 upregulated, 15 downregulated) and 17 transcripts between baseline and discharge samples (11 upregulated, 6 downregulated). In contrast, patients with generalized seizures exhibited a more extensive transcriptomic response, with 74 differentially expressed transcripts between baseline and 4–6 h post-seizure samples (29 upregulated, 45 downregulated) and 70 transcripts between baseline and discharge samples (24 upregulated, 46 downregulated) (FDR-corrected p < 0.05, two-fold change) (Fig. 1c and d). When evaluating persistent transcriptomic changes, we identified one common dysregulated transcript in both the 4–6 h and discharge phases of focal seizures (Fig. 1e). In contrast, a more sustained transcript response was observed following generalized seizures, with 30 transcripts consistently dysregulated at both the 4–6 h and discharge time points compared to baseline (Fig. 1f).
Fig. 1
Temporal dynamics of differential transcript expression in focal and generalized seizures paired with baseline. a Bar graph of 50,081 novel transcripts within the GRCh38 genome. A total of 67.7% were associated with novel genes, and 32.3% with known genes. b Pie chart illustrates the categorization of novel transcripts among which 3191 novel isoforms from novel genes with no overlap with the reference genome. c Line graph of the mean transcript expression of differential expressed transcripts, normalized to the baseline value (100%). Data are separated based on the transcript lists of differentially expressed transcripts for each condition relative to the baseline samples. d Bar plots displayed the dysregulated transcripts (up and down) within the seizure groups with respect to baseline. The number of transcripts increase are represented in purple and decrease in dark green in generalized seizure. Interestingly, an increased transcriptional response was evident shortly after the focal seizure, while in generalized seizures, a more profound transcriptional response occurred at 4–6 h post-seizure but persisted till discharge. e Pie chart depicts consistent dysregulation of one transcript in focal seizure and f 30 transcripts in generalized seizure, indicating distinct temporal transcriptional responses and underlying molecular mechanisms between seizure types
Differential Temporal Expression of Transcripts Following Focal or Generalized SeizuresWe further analyzed transcript expression in patient samples of generalized and focal seizures at baseline, 4–6 h post-seizure, and at discharge (Table 1). The analysis showed 124 transcripts are significantly different between generalized and focal seizure patients at baseline, 42 at 4–6 h post-seizure, and 37 at discharge (FDR-corrected p < 0.05, 2-FC) (Fig. 2c). Interestingly only one transcript overlaps across the distinct time points emphasizing the dynamic nature of seizure-associated transcriptomic profiles (Fig. 2d). The distinct separation between conditions was discernible between focal and generalized seizures in PCA plots of the 4–6 h and discharge samples (24% and 21% variance, respectively) (Fig. 2e).
Fig. 2
Characterization of novel transcripts and differential transcript expression in seizure patients. a Volcano plot showing differentially expressed transcript between focal and generalized seizure at baseline, 4–6 h post-seizure and discharge [black dots—not significant; blue dots—upregulated; red dots—downregulated; orange dots—significant for p value < 0.05 as well as twofold change]. b Venn diagram showing the unique transcripts in following seizure across time points. Note the lack of overlap between these clinical conditions suggesting different sets of transcripts show differential expression over time. c Principal component analysis (PCA) showing a distinct separation between focal and generalized seizures
Differential Transcript Usage Identified Across Seizure TypesWe assessed the differential use of transcript isoforms (isoform switching) using the R package IsoformSwitchAnalyzeR (|dIF|> 0.1) [18]. Isoform switching between focal and generalized seizure revealed 2748 isoforms linked to 2689 switching events in 1249 unique genes. Comparing post-seizure time points to baseline, CORO1C (q = 4.18 × 10⁻32) and ZBTB44 (q = 1.47 × 10⁻20) emerged as top switches in focal seizures (Fig. 3a), while SNHG1 (q = 3.26 × 10⁻54) and RPS17 (q = 2.14 × 10⁻32) were most significant in generalized seizures (Fig. 3b). The highest number of unique switching events was observed in generalized seizure (4–6 h post-seizure, 31.7%; discharge, 26.8%) as compared to focal seizure (4–6 h post-seizure, 5.7%; discharge, 3.0%). However, we also observed small but significant overlaps of common isoform switches following both seizure types (Fig. 3c).
Fig. 3
Top alternatively spliced genes following seizure type compared to baseline, based on q-values. a Focal seizures: CORO1 C (baseline vs. 4–6 h) and ZBTB44 (baseline vs. discharge) were identified as the top-ranked isoforms. b Generalized seizures: SNHG1 (baseline vs. 4–6 h) and RPS17 (baseline vs. discharge) emerged as the top-ranked isoforms. c Unique isoform switching events relative to baseline revealed that focal seizures exhibited 48 distinct switches at 4–6 h post-seizure and 25 at discharge, whereas generalized seizures showed 268 unique switches at 4–6 h and 227 at discharge, highlighting distinct molecular responses between the two seizure types
We then focused on genes associated with significant differential isoform fractions across time points. We show examples of FECH, PPP1R18, and S100A9 as the topmost alternatively spliced genes between focal and generalized seizures in baseline, 4–6 h post-seizure, and discharge samples, respectively. Notably, these genes showed significant isoform changes without differential gene expression (Suppl. Figure 2). Additionally, FECH and S100A9 comprise novel transcripts (MSTRG.27408.6, MSTRG.3657.4, and MSTRG.3657.8) that exhibit differential usage between focal and generalized seizures (Suppl. Figures 2a and c).
The consequence of these switching events on transcript function was then assed. We found a combination of alternative splicing (AS), alternative transcription starts sites (aTSS), and alternative transcription termination sites (aTTS) collectively influenced isoform switching. Specifically, 25.03% (505 genes) of isoform switching events were attributed to the interaction among these mechanisms (Fig. 4a). We noticed a significant (FDR-corrected p < 0.05) increase in alternative transcript termination site (aTTS) (Fig. 4b). These findings suggest that alterations in alternative splicing occur following focal and generalized seizure events.
Fig. 4
Mechanism and functional consequences of isoform switching in focal and generalized seizures a Isoform switching mechanisms involved alternative splicing (AS), alternative transcription starts sites (aTSS), and alternative transcription termination sites (aTTS). b Functional consequences of isoform switching includes alternative transcription starts sites (aTSS) gain as significant splicing event in discharge group
Gene Ontology of Differential Transcript Usage Following SeizuresTo understand the functional implications of genes undergoing switching events, we conducted gene ontology and pathway analyses. We performed functional enrichment analysis of differential transcript usage genes within the seizure groups. Focal seizure group enriched with acute inflammatory and dysregulation of cellular processes, especially protein synthesis and nucleotide metabolism. At discharge, these pathways shift to protein ubiquitination and apoptotic pathway (Fig. 5a and b). Conversely following generalized seizure, pathway analysis shows enhanced cellular activity and metabolic turnover which converge to immune response regulation at discharge (Fig. 5c and d). Notably, following focal seizures, the splicing events do not appear to converge around specific functional pathways. In contrast, following generalized seizures, we observe enrichment in distinct functional pathways.
Fig. 5
Gene ontology and pathway analysis of differential transcript usage genes following seizure types paired with baseline. Focal seizure: a At 4–6 h post-seizure, pathways related to acute inflammatory responses and dysregulation of cellular processes were prominent. b In the discharge group, enrichment shifts to protein ubiquitination and apoptotic pathways. Generalized seizure: c cellular activity and metabolic turnover were increased at 4–6 h post-seizure which d transitioning to immune response regulation by discharge. In contrast to focal seizures, generalized seizures show enrichment in distinct functional pathways, indicating a more coordinated molecular response
We further observed significant enrichment across time points of focal and generalized seizure. In 4–6 h post-seizure group, significant activity is related to the regulation of mRNA splicing and ribonucleoprotein complex assembly (Suppl. Figure 3a) while in discharge group, the focus shifts to immune-related processes, such as lymphocyte activation, cytokine production, and regulation of the immune response (Suppl. Figure 3b). This implies that the immediate post-seizure period is characterized by intense cellular and molecular adjustments, while the later stage involves a significant immune system response.
Differential Transcript Usage Analysis for Seizure Sub-TypeWe further refined signatures associated with sub-type of focal (FIA and FTC) and generalized seizure (GTC) across time points. We identified significant intron retention loss in FIA patients and alternative transcript termination site gain events in GTC patients at baseline and 4–6 h post-seizure, respectively (Fig. 6a). However, intron retention gains with loss of multiple exons skipping events were noticed at discharge in GTC vs. FTC patients (Fig. 6b). We prioritized the top-ranked genes and identified BCL2 (baseline), EIF4G2 (4–6 h post-seizure), and TPI1 (discharge) on FIA vs. GTC comparison and S100A9 (baseline), RAB18 (4–6 h post-seizure), and RN7SL1 (discharge) on FTC vs. GTC comparison. Noticeably, RNF216 was particularly significant in baseline FIA and FTC compared to GTC, with notable NMD-sensitive isoforms (Fig. 6c). These preliminary data suggest patterns of isoform splicing are different following different sub-types of seizure.
Fig. 6
Identification of signatures associated with seizure sub-types across time points. a Statistically significant functional alterations are observed in comparisons including baseline FIA vs. GTC (intron retention gain), 4–6 h FIA vs. GTC (alternative transcription termination sites), and b discharge FTC vs. GTC (intron retention gain and multiple exon skipping). c RNF216 exhibits common isoform switching in baseline FIA and FTC compared to GTC. FIA, focal with impaired awareness; FTC, focal seizures time course; GTC, generalized tonic clonic]
ML-Based Seizure Prediction Highlights the Difference Between Focal and Generalized SeizuresWe investigated whether ML prediction modeling could be used to identify patients who have a specific seizure sub-type vs. a non-seizure (baseline) sample. The goal of the modeling was to determine whether a focal seizure would be identified as a generalized seizure and vice-versa. Classifier lists of transcripts were initially established following differential transcript expression analysis to identify baseline vs 4–6 h differential transcript expression following focal seizure (Fig. 1c). We first created a training dataset of focal seizure samples, consisting of baseline samples from patients with EEG-confirmed focal seizure (N = 21) and 4–6 h post-focal seizure samples (train = 20). The testing set consisted of baseline and 4–6 h samples from patients with EEG-verified generalized seizure (N = 6 of each). Using the training data (focal), we trained classifiers using six algorithms from the caret package, as depicted in Fig. 7a. The model was trained on baseline versus 4–6 h post-seizure data from patients who had experienced a focal seizure (21 patients; two samples each), with each subjected to ten-fold cross-validation. The AUC-ROC, sensitivity, and specificity of cross validation testing are given in Fig. 7a and b; random forest, Naïve Bayes, and radial SVM models exhibited the highest performance during training (mean AUC-ROC values ranging from 0.95 to 1.0) (Fig. 7a). The training model when applied to the training data achieved 100% accuracy as expected. These models were subsequently tested on baseline and 4–6 h generalized seizure data (six patients; two samples each). The models classified 11 samples as no focal seizure (N) (Fig. 7c). A second model was trained only the generalized seizure data using the same algorithms (generalized train). Based on AUC-ROC analysis, the multivariate adaptive regression splines (mars) model performed best, achieving an AUC-ROC value of 1.0 (Fig. 7d). The performance evaluation of these classifiers is shown in Fig. 7e. This model was then tested on the focal seizure data, and it classified all samples as no generalized seizure (N), correctly identifying focal seizure data as not generalized seizures (Fig. 7f). These data suggest ML models based on focal seizure data do not misclassify generalized seizure data and vice versa.
Fig. 7
ML-based modeling to distinguish between focal and generalized seizure. a Classification models were trained using six algorithms on baseline versus 4–6 h post-seizure data from focal seizure patients. Random forest, Naïve Bayes, and radial SVM exhibited the highest performance with AUC-ROC values ranging from 0.95 to 1.0. These models achieved 100% accuracy on training data and correctly classified generalized seizure data without misclassification. b Box plots summarize the specificity, AUC-ROC, and sensitivity metrics for models trained on focal seizure data, showing the distribution of values across different models. c A confusion metric presents the classification results when the focal seizure-trained model was tested on the generalized seizure dataset, highlighting actual versus predicted classifications. d A second model was trained on generalized seizure data using the same algorithms, with MARS performing best, achieving an AUC-ROC value of 1.0. e Box plots depicting performance variation across different models while f confusion metrics summarized 100% accuracy, correctly identifying all focal seizure data as not generalized seizures. (model abbreviations: glm, generalized linear model; rf, random forest, NB, naïve Bayes; mars, multivariate adaptive regression splines, svm, support vector machine, DT, decision tree)
We also performed prediction modeling using differentially expressed transcripts between baseline and discharge samples following EEG-verified focal seizures (Fig. 1c). Using the same models and parameters as for the 4–6 h data, the random forest algorithm showed good training performance on the focal seizure dataset. However, the model misclassified one baseline sample as a focal seizure and two generalized seizures as focal seizures (Suppl. Figure 4a). Similarly, training on the generalized seizure dataset yielded acceptable training performance (RF accuracy = 0.9), but the model struggled to correctly classify generalized seizures, misclassifying seven focal seizures and 13 baseline samples as generalized seizures (Suppl. Figure 4b). These findings suggest that the 4–6 h data may be more predictive, or that the model requires a larger dataset for improved classification.
Comments (0)