
Remember me

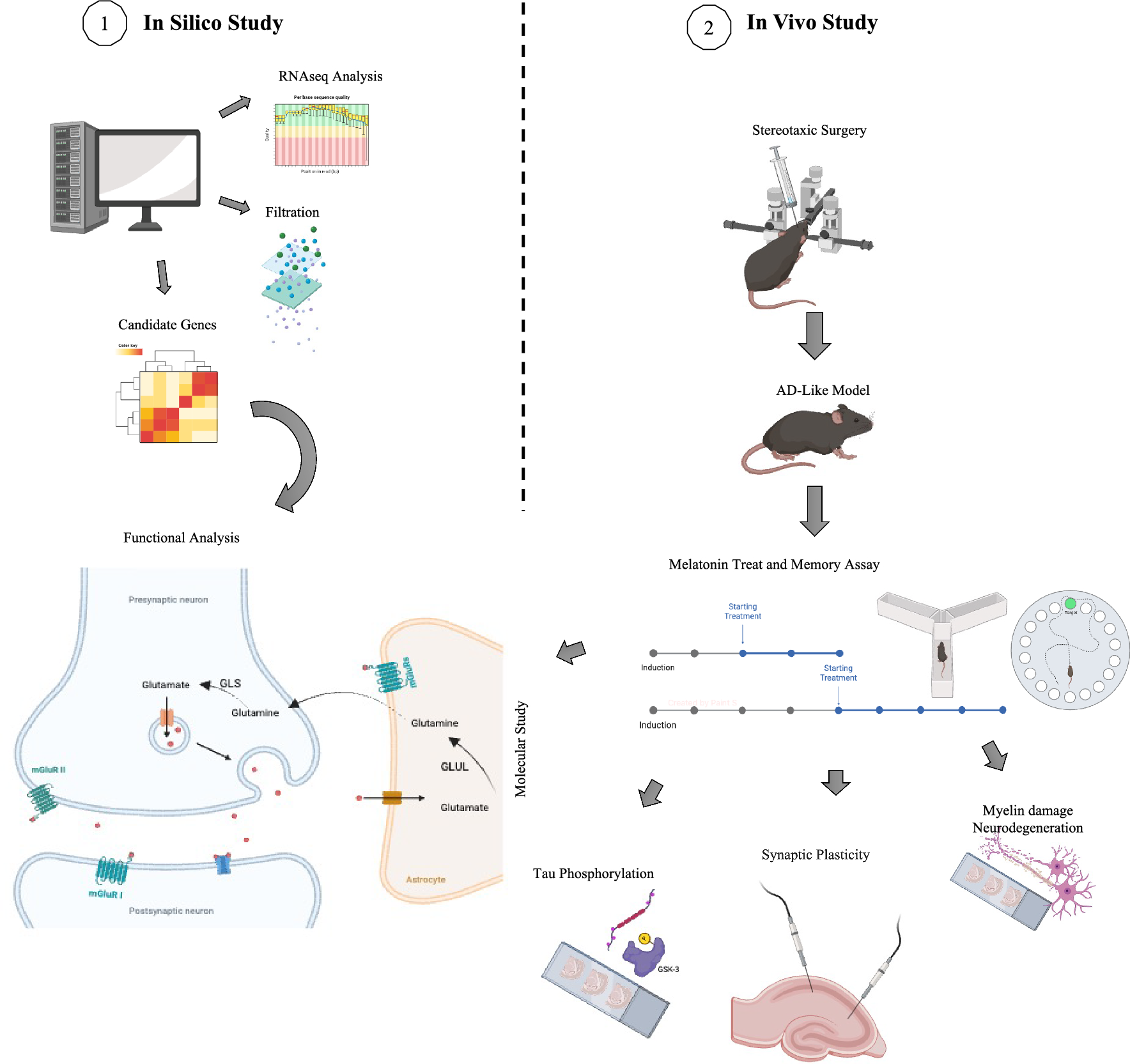


In silico analyses were employed to uncover potential molecular mechanisms through which melatonin might ameliorate cognitive impairments. For RNA-seq analysis, data from the hippocampal tissues of AD patients were obtained from the datasets GSE67333, GSE184942, and GSE236562. These datasets were selected based on similar mean ages and disease stages (ABC Scoring system) (Table 3).
Table 3 Demographic data of RNA sequencing datasetsRNA-seq analysis generated the Gene Count Table, and Batch Effect Correction was performed using the ComBat-Seq model [23]. Differential Gene Expression (DEG) analysis was conducted using the DeSeq2 package in R. Target genes were filtered from the Mouse Genome Database (MGD) and the Gene Ontology (GO) Database, focusing on learning, memory, neuroplasticity, and behavior regulation genes. Given the significant role of GPCR receptors in neurodegenerative disorders, a GPCR gene list was sourced from the HUGO Gene Nomenclature Committee (HGNC) database[24].
With criteria of −0.5 ≥ log2FC ≥ 0.5 and 0.05 ≥ p-value, a Volcano plot was generated for candidate genes (Fig. 1A) (Additional file 2). Principal Component Analysis (PCA) illustrated gene expression differences between control and AD samples (Fig. 1B). The glutamatergic system and mGluR receptors were selected for molecular investigation (Fig. 1C).
Fig. 1
Transcriptome analysis and gene expression changes in the hippocampus of AD patients. A Volcano plot depicting gene expression changes. The x-axis represents log2 fold change (log2FC), and the y-axis represents the p-value. Genes with −0.5 ≤ log2FC ≤ 0.5 and p-value ≥ 0.05 are considered significant. Blue dots on the right indicate increased expression, while dots on the left indicate decreased expression in AD samples. Red dots represent GPCR genes outside the specified range. This plot was generated using the ggplot2 package in R. B Principal Component Analysis (PCA) graph illustrating differences between control and AD samples based on gene expression. C Heatmap showing significant expression changes in genes involved in the glutamatergic system. Each row on the Y-axis represents a single gene, and dendrograms represent genes with similar expressions. The X-axis represents all samples, and the dendrogram clusters control and patient samples. The color gradient from blue to red indicates low to high expression levels. This plot was created using the p-heatmap package in R
The therapeutic effect of melatonin on working memory impairment in AD-like animalsIn the first experiment, we investigated whether melatonin had a therapeutic effect in animals with mild memory impairment, AD-like model mice received melatonin at two weeks post-cis-P tau injection, and the working memory performance was assessed using the Y-Maze test. A two-way ANOVA was performed to assess the significance of differences in working memory performance across study groups and time points. The results demonstrated a statistically significant difference among the study groups (F (2,33) = 251.5, p < 0.0001) and across the evaluated time points (F (3,99) = 450, p < 0.0001). Melatonin facilitated working memory in treated AD-like mice (Fig. 2A, AD-Melatonin). As Fig. 2A shows, the spontaneous alternation was significantly increased in these animals after 2 weeks of treatment compared to the AD-like group (t = 20.88, p < 0.001). Interestingly, there was no significant difference in working memory between treated animals and the control group in 4th week (Fig. 2A). In contrast, untreated AD-like mice (Fig. 2A, AD) exhibited progressive memory impairment (t = 29.55, p < 0.0001).
Fig. 2
Improvement of working memory impairment due to melatonin treatment in AD-like animal model. A Working memory assessment pre- and post-melatonin treatment when the melatonin injection (10 mg/kg) was initiated at the 2nd week post-induction. B Working memory assessment pre- and post-melatonin treatment when the melatonin (10 mg/kg) was initiated at the 4th week post-induction. The results represent the average performance of 12 samples per group, indicating observed trends in working memory improvement. *** p < 0.001 compared to control group, ### p < 0.001 compared to AD group
In the second experiment, the therapeutic action of melatonin was evaluated after establishing a severe memory impairment in animals. Melatonin treatment was started at the fourth-week post-cis p-tau injection and continued for 4 weeks. The results of two-way ANOVA demonstrated a statistically significant difference among the study groups (F (2,33) = 679.7, p < 0.0001) and across the evaluated time points (F (7,245) = 49.79, p < 0.0001). The obtained results showed the improving effect of melatonin on working memory, so the spontaneous alternation was increased in the AD-melatonin group compared to the AD group (t = 11.45, p < 0.0001). The significant increase in spontaneous alternation was started after the 2nd week of melatonin injection (i.e., at the 6th to 8th weeks of the experiment; Fig. 2B). However, the memory impairment remained in these animals and did not reach the control values (Fig. 2B) (t = 35.38, p < 0.0001). These findings suggested that earlier melatonin intervention yielded better cognitive improvement.
The therapeutic effect of melatonin on spatial memory and learning impairment in AD-like animalsSpatial learning impairments were evident in the AD group (Figs. 3 and 4), as demonstrated by prolonged primary (Figs. 3B and 4A) (F (1, 88) = 1848, p < 0.0001 and F (1, 88) = 4195, p < 0.0001, AD 4 weeks and AD 8 weeks respectively) and total latencies (Figs. 3C and 4B) (F (1, 88) = 1499, p < 0.0001 F (1, 88) = 3645, p < 0.0001, respectively) and increased primary (Fig. 3D and 4C) F (1, 88) = 778.0, p < 0.0001 and F (1, 88) = 2612, p < 0.0001, respectively) and total errors (Figs. 3E and 4D) (F (1, 88) = 7337, p < 0.0001 and F (1, 88) = 4168, p < 0.0001, respectively) across training days. In contrast, the AD-Melatonin group showed significant improvement in all parameters (For primary latencies: F (1, 64) = 765.6, p < 0.0001 and F (1, 64) = 898.7, p < 0.0001, AD 4 weeks-Melatonin and AD 8 weeks-Melatonin respectively. For total latencies: F (1, 64) = 758.7, p < 0.0001, and F (1, 64) = 182.0, p < 0.0001, respectively. For primary error: F (1, 64) = 409.6, p < 0.0001, F (1, 64) = 264.4, and for total error: F (1, 64) = 3744, p < 0.0001 and F (1, 64) = 505.6, p < 0.0001, respectively), achieving performance comparable to controls when melatonin treatment was initiated two weeks post-Cis-Phospho Tau injection. The strategy to find the goal box was also assessed (Figs. 3F and 4E). The random strategy shows the deficiency in animal’s learning and memory. In serial strategy, the animal's learning increases, and the subject can find the goal box using its memory. Delayed treatment (initiated four weeks post-injection) improved learning metrics but did not fully restore performance to control levels.
Fig. 3
Improvement of spatial memory and learning impairment due to melatonin treatment initiated at the 2nd-week post-induction. A Diagram illustrating the circular Barnes maze with 20 equally spaced holes, including the goal sector (GS) and non-goal sector (NGS). BPrimary latency to locate the goal hole. C: Total latency to enter the goal box. D Primary errors during exploration. E Total errors across training days. F Search strategies (direct, serial, and random) adopted by animals across training days. G Goal sector exploration frequency. H Non-goal sector exploration frequency. I Goal sector preference (goal/non-goal exploration ratio). The results represent the average performance of 12 samples per group, and data are presented as mean ± SEM. Statistical significance: *** p < 0.001 compared to the control group, ### p < 0.001 compared to the AD group
Fig. 4
Reduced improvement in spatial memory and learning impairment due to delayed melatonin treatment initiated at the 4th-week post-induction. A Primary latency to locate the goal hole. B Total latency to enter the goal box. C: Primary errors during exploration. D Total errors across training days. E Search strategies (direct, serial, and random) adopted by animals across training days. F Goal sector exploration frequency. G Non-goal sector exploration frequency. H Goal sector preference (goal/non-goal exploration ratio). The results represent the average performance of 12 samples per group, and data are presented as mean ± SEM. Statistical significance: *** p < 0.001 compared to the control group, ### p < 0.001 compared to the AD group
The probe test revealed significant memory and learning deficits in the AD group (Figs. 3G–I, and 4F–H), as indicated by reduced goal sector (GS) preference (t = 22.28, p < 0.0001 and t = 22.04, p < 0.0001, AD 4 weeks-Melatonin and AD 8 weeks-Melatonin respectively) and increased non-goal (NGS) exploration (t = 9.225, p < 0.0001 and t = 14.90, p < 0.0001, AD 4 weeks-Melatonin and AD 8 weeks-Melatonin respectively). Melatonin treatment initiated two weeks post-injection restored these parameters (for GS: t = 10.27, p < 0.0001 and for NGS: t = 6.323, p < 0.0001, compart to AD group) to control levels. Delayed treatment resulted in partial recovery, with significant improvement compared to the AD group but not reaching control levels.
Melatonin treatment reduces myelin sheath degradation and neuronal death in the hippocampus of AD-Like modelRecent studies have indicated that cognitive impairments may reflect the degradation and deficiency of myelin in the brains of animal models [25]. Therefore, Luxol Fast Blue.
Staining was used to examine myelin changes and the effect of melatonin treatment in AD-like model animals (Fig. 5A). Results revealed significant thinning of the myelin sheath in AD-like model groups, more pronounced at the 8th week (Fig. 5B) (t = 8.586, p = 0.001, and t = 18.13, p = 0.0001, respectively). Melatonin treatment initiated in the 2nd week post-induction significantly increased Luxol Fast Blue intensity (t = 3.707, p = 0.01), although it did not restore levels to those of the control group. However, melatonin treatment initiated at the 4th-week post-induction did not improve myelin damage (t = 1.751, p > 0.05).
Fig. 5
The effect of melatonin on reducing CA1 hippocampal tissue damage in the AD-like model. A Luxol Fast Blue (LFB) staining of the CA1 hippocampal region, illustrating the extent of myelin preservation and demyelination across different experimental groups. LFB is a histological stain that specifically binds to lipoproteins in the myelin sheath, allowing visualization of myelin integrity. Intact myelin appears deep blue, whereas demyelinated or damaged regions show lighter staining or loss of staining. This technique is particularly relevant in neurodegenerative conditions such as AD, where white matter integrity is progressively compromised. B Graph showing the quantitative intensity of LFB staining, representing the average percentage of myelin staining in samples from each group. This provides a comparative assessment of myelin loss and its potential restoration following melatonin treatment. C Nissl staining indicating neuronal death, with arrows pointing to degenerated or pyknotic cells in the CA1 region. C: Nissl staining indicating neuronal death, with arrows pointing to dead cells. D Quantitative analysis of the percentage of dead cells in the CA1 region, with data averaged across the samples in each group. The tissue samples displayed in the histological images include Control animals, an AD-like model with 4 weeks of disorder (AD 4W), and model under melatonin treatment (AD 4W-Melatonin), an AD-like model with 8 weeks of disorder (AD 4W), and model under melatonin treatment (AD 8W-Melatonin). The groups in the graphs are presented as Control animals, AD-like models (AD) with 4 weeks (4W) and 8 weeks (8W) of disorder, and models under melatonin treatment (AD-Melatonin) starting from the second-week post-induction (4W) and starting from the fourth-week post-induction (8W). The symbol * indicates statistically significant differences compared to the Control, and the symbol # compared to the model animals without treatment
Previous reports have shown that neuronal death in the hippocampus can lead to cognitive impairments; thus, neuronal death was assessed using Nissl staining in the CA1 region of the hippocampus across the study groups. Nissl staining showed increased neuronal death in AD-like models (Fig. 5C, D) (t = 10.46, p = 0.0005), with a greater extent of death at the 8th week (t = 9.889, p = 0.0006). Melatonin treatment significantly reduced neuronal death (t = 10.46, p = 0.01 and t = 4.331, p < 0.05, respectively), confirming that earlier intervention was more effective.
Positive impact of melatonin treatment on pathogenic changes in Gls1 and Glul protein levels in the AD-Like animal modelConsidering the importance of glutamate in cognitive activities such as memory, and the findings from the in-silico study, we examined the changes in transcriptomic (Additional file 3) and protein levels of two key enzymes involved in glutamate homeostasis, Gls1 and Glul, in hippocampal samples from animal model groups treated with melatonin.
Western blot analysis revealed increased Gls1 protein levels in AD-like models (Fig. 6A–C). Descriptive analysis showed that melatonin treatment initiated in the 2nd-week post-induction reduced Gls1-65 KDa protein levels, although they did not return to control levels. The 58 KDa isoform of Gls1 remained mostly unchanged in the early stages but showed a reduction with later treatment. Similarly, Glul protein levels (Fig. 6D), which were decreased in AD-like models, demonstrated partial restoration following melatonin treatment.
Fig. 6
The effect of melatonin treatment on Gls1, Glul, GSK3, and phosphorylated GSK3 levels in AD-like models. A Western blot for protein quantification in study animals, including Control animals, AD-like model at 4 weeks (AD 4W) and 8 weeks (AD 8W) post-induction, and under melatonin treatment initiated at two different time points. B Gls1-65 KDa protein levels across the experimental groups. C Gls1-58 KDa protein levels, illustrating the relative changes among the groups. D Glul protein levels showing trends of variation in response to melatonin treatment. E GSK3α protein levels across the groups. F GSK3β protein levels, highlighting differences between treated and untreated groups. G Phosphorylated GSK3α levels, indicating changes in kinase activity under melatonin treatment. H Phosphorylated GSK3β levels, showing the effect of treatment on kinase activity. The results were derived from pooled samples, with 5 samples pooled per group. Due to pooling, statistical analysis was not performed, and the data are presented descriptively
In summary, melatonin treatment had a positive effect in modulating pathogenic changes in Gls1 and Glul protein levels, which are critical for maintaining glutamate homeostasis and cognitive function. The effectiveness of melatonin was more pronounced when treatment started earlier, highlighting the importance of early intervention in neurodegenerative conditions.
Positive impact of melatonin treatment on gene expression changes in mGluR receptors observed in AD-Like animal modelsOur literature review and bioinformatics analyses indicated that metabotropic glutamate receptors (mGluRs) play crucial roles in various CNS functions such as learning, memory, and synaptic plasticity. Therefore, we investigated these GPCRs as functional mediators of cognitive impairment in our AD-like animal models and assessed the effects of melatonin treatment on their expression patterns. mGluRs are widely distributed throughout the CNS, responsible for synaptic modulation, and are classified into three subgroups based on their signaling pathways. Group I mGluRs include mGluR1 and mGluR5 (gene names: Grm1, Grm5), Group II includes mGluR2 and mGluR3 (gene names: Grm2, Grm3), and Group III includes mGluR4, mGluR6, mGluR7, and mGluR8 (gene names: Grm4, Grm6, Grm7, Grm8). This study examined the transcriptomic changes of seven mGluRs, with the qPCR data heatmap displayed in Fig. 7 reflecting Grm expression levels in the animal model groups studied (Additional file 4).
Fig. 7
Positive effect of melatonin treatment on mGluRs expression in the AD-like models. Heat map created from qPCR data reflecting the expression of Grm genes across the study animal groups. Columns represent the studied animal samples, including Control, AD-like models with 4 weeks of disorder (AD 4W) and those under melatonin treatment (AD 4W Melatonin), AD-like models with 8 weeks of disorder (AD 8W), and models under melatonin treatment (AD 8W-Melatonin). Rows represent different Grm genes, including Grm1 and Grm5 related to the mGluR I group, Grm2, and Grm3 related to the mGluR II group, and Grm4, Grm7, and Grm8 related to mGluR III group. The intensity of color corresponds to the relative expression for each gene calculated using 2^−∆Ct. The color gradient from red to white indicates high to low expression levels, reflecting relative gene expression among the different groups. Results are presented as the average values from independent biological samples (n = 5) within each group
qPCR analysis showed increased Grm1 and Grm5 expression in 4-week AD-like models (Fig. 7) (t = 35.63 and t = 39.76, p < 0.0001), but melatonin treatment reduced their expression significantly (t = 31.48 and t = 37.46, p < 0.0001). Conversely, 8-week models showed decreased expression (t = 8.048 and t = 12.90, p ≤ 0.001), however, melatonin treatment partially restoring levels (t = 4.63, p = 0.0001 and t = 7.92, p ≤ 0.01, respectively), suggesting progressive synaptic and cellular deterioration over time in untreated AD-like conditions.
The expression levels of Grm2 (t = 7.440, p = 0.01; t = 7.96, p = 0.01) and Grm3 (t = 5.47, p = 0.01; t = 23.22, p < 0.0001) were significantly elevated in both the 4-week and 8-week AD-like models compared to controls (Fig. 7, Additional File 4). Early melatonin treatment effectively attenuated these increases, bringing Grm2 and Grm3 expression levels closer to control values (t = 6.656, p = 0.01; t = 6.413, p = 0.01) in the 4-week models. However, in the 8-week models, late melatonin treatment exhibited only partial restorative effects on Grm3, with negligible impact on Grm2, highlighting the critical importance of early intervention. These findings suggest that the initial upregulation of mGluR2 and mGluR3 in the 4-week models may represent a compensatory response to early excitotoxicity and synaptic stress. However, as the disease progresses (8-week models), neuroinflammatory pathways and tau pathology might contribute to receptor desensitization and reduced expression, leading to further synaptic dysfunction.
Grm4 expression increased significantly in 4-week models (t = 7.166, p = 0.01), while Grm7 showed no significant change. Grm8 expression decreased slightly but significantly (t = 3.67, p < 0.05). Melatonin treatment improved Grm7 and Grm8 levels when initiated early (t = 6.46, p = 0.01 and t = 15.22, p = 0.001, respectively).
Overall, our findings demonstrate that melatonin treatment positively impacted the expression of multiple mGluR genes, which are essential for cognitive function and synaptic regulation. Importantly, the effect was more pronounced when treatment was initiated early, reinforcing the significance of timely therapeutic intervention in neurodegenerative conditions.
These results suggest that mGluR dysregulation in AD probably is dynamic and time-dependent, with early alterations likely representing a compensatory mechanism, whereas later reductions may reflect progressive neurodegeneration and receptor desensitization. The ability of melatonin to restore mGluR expression in early-stage models highlights its potential neuroprotective role in modulating glutamatergic signaling, which may contribute to its beneficial effects on cognitive impairment in AD-like conditions.
Melatonin treatment reduces pathogenic phospho-tau levels and GSK3 kinase activity in AD-like animal modelGiven the well-established role of the kinase enzyme GSK3 in tau phosphorylation and the pathobiology of AD, along with its downstream targeting by the receptors studied, we examined the gene expression (Additional file 5), protein levels, and phosphorylation of the two GSK3 enzyme isoforms, GSK3α and GSK3β, in the animal model groups. The activity of these kinase isoforms is indicated by the phosphorylation of tyrosine residue 279 in GSK3α (pTyr279) and tyrosine 216 in GSK3β (pTyr216).
Western blot results indicated increased GSK3α and GSK3β levels in AD-like models. Descriptively, melatonin treatment appeared to reduce both GSK3α and GSK3β protein levels and their phosphorylation status in these models.
One of the targets of GSK3 kinase is the phosphorylation of the threonine 231 residue of the tau protein (pT231-tau). Therefore, to investigate the tissue changes in phosphorylated tau levels under melatonin treatment in the animal models, we performed immunofluorescence staining for pT231-tau (Fig. 8A). Immunofluorescence staining for pT231-tau showed significant increases in both models (Fig. 8B) (t = 15.70, p < 0.0001 and t = 25.90, p < 0.0001, respectively), which melatonin treatment reduced (t = 9.93, p = 0.001 and t = 5.52, p = 0.01, respectively), though not to baseline.
Fig. 8
Melatonin treatment leads to a decrease in tau phosphorylation levels in the AD-Like model. A Immunofluorescence staining for pT231-tau (Green = cis pT231-tau, Blue = DAPI). B Quantitative graph of pT231-tau. Results are presented as the average values from independent biological samples (n = 5) within each group. Groups in the images are Control, AD-like model with 4 weeks of disorder (AD 4W) and under melatonin treatment from the second-week post-induction (AD 4W Melatonin), AD-like model with 8 weeks of disorder (AD 8W) and under melatonin treatment from the fourth-week post-induction (AD 8W Melatonin); and in the graphs as Control animals, AD-like models (AD) with 4 weeks (4W) and 8 weeks (8W) of disorder, and models under melatonin treatment (AD-Melatonin) starting from the second-week post-induction (4W) and starting from the fourth-week post-induction (8W). The symbol * indicates statistically significant differences compared to the Control, and the symbol # compared to model animals without treatment
These findings suggest that melatonin treatment can modulate the pathogenic activity of GSK3 and the phosphorylation of tau protein in an AD-like animal model. The results emphasize the potential of melatonin as a therapeutic agent targeting kinase pathways to mitigate the progression of AD-related pathology, though earlier interventions proved more effective.
Restoring effect of melatonin treatment on LTP generation in AD-like modelFollowing the observed improving effect of melatonin on working memory and changes in mGluR receptors in AD-like model animals, and considering the impact of these receptors on synaptic plasticity, the effect of melatonin was investigated on LTP generation in the AD-like model. Field potential recordings showed a significant decrease in LTP induction (t = 2.459, p < 0.05) and maintenance (t = 3.11, p < 0.01) in AD-like (Fig. 9A and B). Melatonin treatment restored LTP generation when administered in the second week after cis-P tau injection. There was a significant difference in LTP induction (t = 1.146, p < 0.05) and maintenance (t = 2.155, p < 0.01) between AD-melatonin-2nd week and AD groups (Fig. 9). However, melatonin injected at 4th week after cis-P tau injection had no significant effect on LTP (Fig. 9). These results revealed that the restoring effect of melatonin on LTP may observe only in animals with mild memory impairment.
Fig. 9
Improvement of long-term potentiation (LTP) generation by melatonin in the AD-like animal model. A Timeline curve showing the impairment in LTP generation in the AD group that was restored when melatonin was injected in the 2nd week, but not in the 4th week, after pT231-tau injection in the AD-Like animal model. B Percentage of LTP induction and maintenance in different experimental groups animal groups. Each group consisted of 6 samples, with 3 technical replicates per sample. * p < 0.05 compared to the Control group
Comments (0)