
Remember me


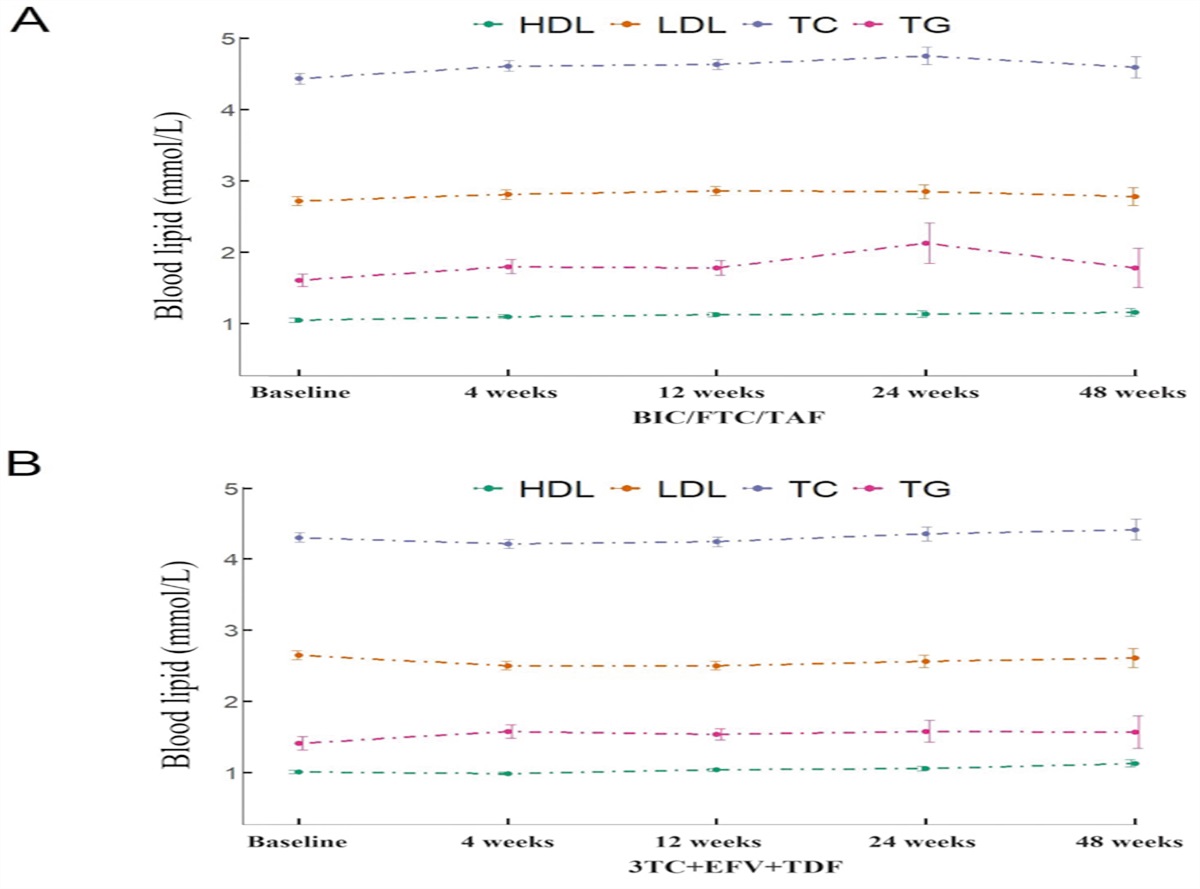
The global population is increasing in terms of growth in size as well as a rise in the ratio of older people to younger ones.[1] There is a growing recognition that aging itself as a natural consequence of living plays a significant role in the development of multiple chronic diseases, including frailty, stroke, arthritis, neurodegeneration, sarcopenia, cancer, vascular disease, renal failure, dementia, diabetes mellitus, osteoporosis, and macular degeneration.[2] Age-related diseases are due to a time-dependent accumulation of cellular damage that causes a progressive decline in cellular and physiological functions, thereby increasing the risk of diseases and death. The effects of aging are not only on the elderly but also on those of younger ages. Particularly, the increased chronic oxidative stress and inflammation have been shown to accelerate aging even in the younger ages.[2] Thus, age-related diseases represent a substantial challenge to health care and socioeconomics worldwide. Studies on targeting fundamental aging mechanisms have been critical in the gerontology community.[3] Environmental factors, including exposure to pollutants, diet, and sedentary lifestyle, largely influence human health and lifespan by triggering age-associated cellular and molecular events.[4] Aryl hydrocarbon receptor (AhR) is a cytoplasmic receptor and mainly expressed at barrier surfaces, including the skin, respiratory tract, and gastrointestinal tract, linking environmental, microbial, and metabolic cues to biological and pathophysiological changes through regulating complex transcriptional programs.[5–8] In recent years, mounting evidence has revealed that AhR plays an important role in regulating environmental factor-triggered age-associated phenotypes. The human lung is one of the important organs highly impacted by aging and has the largest surface area exposed to environmental pollutants and biological, immunological, and xenobiotic stress.[9,10] Recent advances indicate that asthma in the elderly population represents a unique phenotype, highlighting that age is a significant risk factor for the inflammatory features, exacerbations, and poor response to treatment in the elderly population with asthma.[11]
In this review, we discuss the literature pertaining to the hallmarks of aging, including mitochondrial dysfunction, cellular senescence, autophagy/mitophagy, epigenetic alterations, inflammation, and microbiome disturbance. We propose AhR as a ligand-activated transcription factor linking environment to immunity regulating hallmarks of aging. Finally, we evaluate the potential role of AhR in regulating asthma phenotypes in the elderly population linked to the aging process.
Mechanisms of Aging ProcessesAging appears to be a multi-dimensional, irreversible accumulation of physical, environmental, and social changes with a progressive decline in the physiological functions of an organism.[12] Aging can be accelerated by environmental exposure-induced chronic inflammation and subsequent excessive oxidative stress. Specifically, environmental exposure-induced reactive oxygen species (ROS) within mitochondria can damage the mitochondria, which in turn leads to the overproduction of ROS that causes further damage, and thereby aging. However, studies on the cellular and molecular mechanisms of aging processes associated with age-related diseases are limited. Understanding the mechanisms of the aging process is therefore crucial for the prevention or treatment of multiple age-related diseases. Notably, the current research on the biology of aging is guided by the cellular and molecular hallmarks of aging defined by López-Otín et al.[13] These hallmarks of aging include genomic instability, telomere attrition, epigenetic alterations, mitochondrial dysfunction, loss of proteostasis, deregulated nutrient-sensing, cellular senescence, stem cell exhaustion, and altered intercellular communication. Recent advances suggest the role of many new hallmarks of aging, including autophagy, microbiome disturbance, altered mechanical properties, splicing dysregulation, and inflammation.[13,14] The updated hallmarks of aging may provide a better guide and advanced research on the biology of aging, as illustrated in Figure 1.
 Figure 1:
Figure 1: Updated hallmarks of aging.
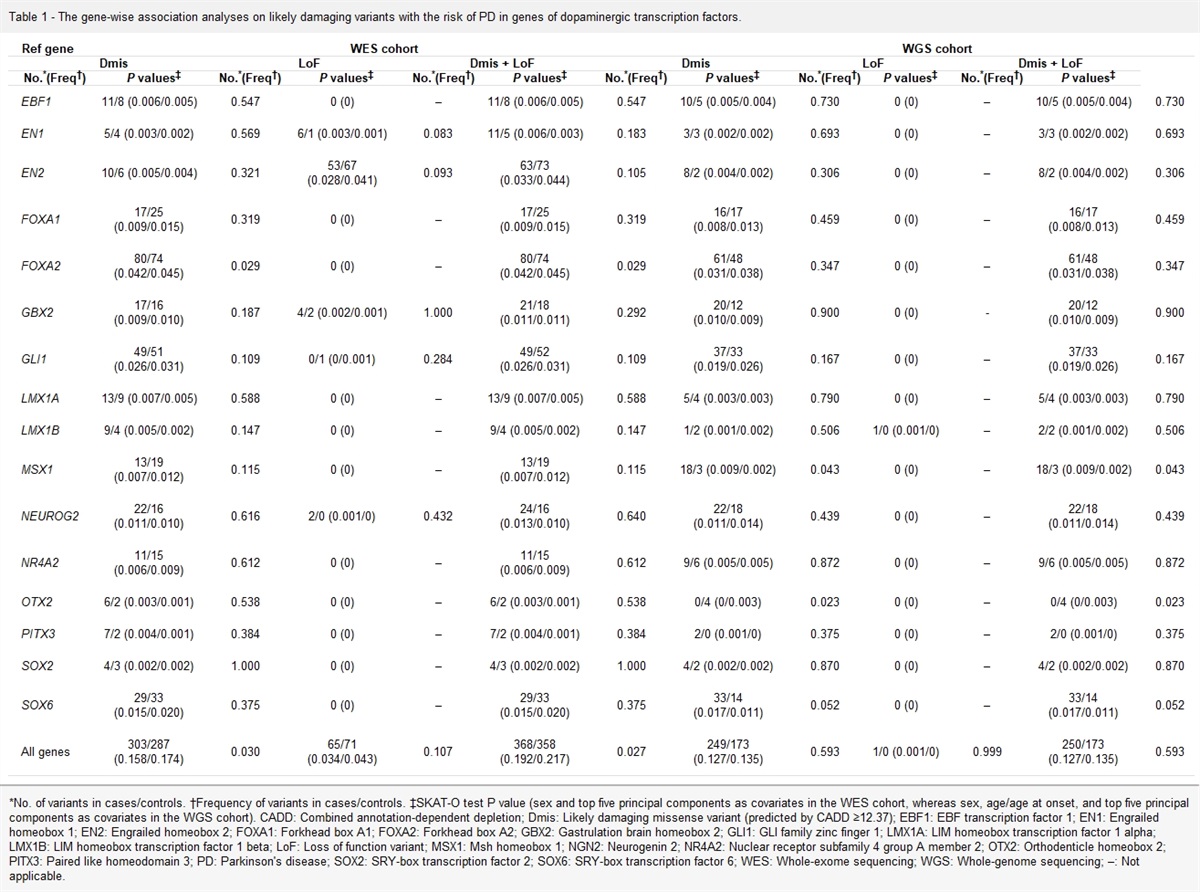
Mitochondrial dysfunctionMitochondrial dysfunction has been considered a hallmark of environmental injury.[4] The major feature of mitochondrial dysfunction in aging tissues is a decrease in respiratory capacity per mitochondrion and in mitochondrial membrane potential (MMP).[15] During mitochondrial dysfunction, low MMP is mainly linked to the overproduction of ROS.[16] Furthermore, mitochondrial biogenesis declines with age, reducing adenosine triphosphate (ATP) generation and increasing electron transport. These age-dependent abnormalities in mitochondrial biogenesis can contribute to the impairment and weakening of mitochondrial function. With advanced age, mitochondrial DNA (mtDNA) volume, integrity, and functionality can decrease, and mitophagy, an autophagy process that removes dysfunctional mitochondria due to accumulation of oxidative damage induced by ROS, is inhibited.[17] ROS are critical in signaling cellular stress that initially compensates for age-related deterioration, which seems beneficial for mitochondrial homeostasis.[18] As age progresses, excessive ROS can exacerbate age-related mitochondrial oxidative damage, implying ROS’s role in aging.[19,20] Several mechanisms cause mitochondrial dysfunction, including mtDNA mutations (genomic instability), mitochondrial turnover as measured by the ratio of mitochondrial biogenesis and mitophagy associated with fusion and fission, nutrient signaling through mammalian target of rapamycin (mTOR) regulated by the mitochondrial sirtuin 3 (SIRT3) and SIRT5, nicotinamide adenine dinucleotide/nicotinamide adenine dinucleotide hydrogen (NAD+/NADH) imbalance, and Ca2+ fluxes resulting in mitochondrial Ca2+ overload [Figure 2].[15] Of these, mtDNA shows an increased susceptibility to ROS, leading to severe oxidative damage.[21] The mutation rate of mtDNA is 10–17 times higher than that of a nuclear genome (nDNA) because of the lack of histones protection and efficient DNA repair mechanism.[22] mtDNA mutations can cause mtDNA damage, and the accumulation of mtDNA damage with age and impaired mtDNA repair can lead to mitochondrial dysfunction.[23] Thus, mtDNA is a major risk factor that contributes to mitochondrial dysfunction and age-associated multisystemic diseases.
 Figure 2:
Figure 2: Mechanisms that can cause mitochondrial dysfunction. These include mitochondrial Ca2+ overload, mitochondrial turnover, nutrient signaling, genomic instability, and NAD+/NADH imbalance. NAD+: Nicotinamide adenine dinucleotide; NADH: Nicotinamide adenine dinucleotide hydrogen.
Dysregulation of nutrient cellular signaling is also one of the mechanisms contributing to mitochondrial dysfunction through mTOR, which is modified by the mitochondrial SIRT3 and SIRT5.[15] Particularly, excessive cellular nutrients activate the mTOR pathway and induce anabolic processes, and cause inhibition of autophagy. In contrast, a lack of cellular nutrients activates mitochondrial SIRT3 and SIRT5, resulting in mitochondrial biogenesis and increased autophagy.[24] The imbalance of the NAD+/NADH ratio also causes cellular and mitochondrial dysfunction during aging.[25] NAD+ is a cofactor in several oxidation reduction pathways and a substrate for many redox reactions. While NADH is produced by glycolysis and tricarboxylic acid (TCA) cycle in the mitochondria, NAD+ as a cofactor is required for the production of ATP and maintenance of MMP. Thus, the optimal ratio of NAD+/NADH is required for normal cellular reactions and mitochondrial metabolism, and disruption of NAD+/NADH ratio can affect mitochondrial function, and subsequently aging.
Cellular senescenceCellular senescence is defined as a state of permanent cell cycle arrest induced by several factors, including aging, oxidative stress, DNA damage, mitochondrial dysfunction,[26] epigenetic modifications,[27] and telomere shortening.[28] Senescence arrest occurs in the G1 phase of the cell cycle, distinguishing it from G0-arrested quiescent cells, and is mediated by cyclin-dependent kinase inhibitors (CDKis) (e.g., p21CIP1, p16INK4a) and is also dependent on the tumor protein p53 (TP53) and retinoblastoma protein (pRB) tumor suppressor pathways.[29] Senescent cells can secrete plethoric pro-inflammatory cytokines, chemokines, angiogenic factors, growth modulators, and matrix metalloproteinases (MMPs), termed the senescent associated secretory phenotype (SASP). The SASP can create a feedback loop, further promoting senescence in neighboring cells and contributing to tissue dysfunction.[30] Senescent cells also show increased rates of mitochondrial metabolic activity, including the TCA cycle, oxidative phosphorylation (OXPHOS), and glycolytic pathways, together with increased adenosine monophosphate (AMP) and adenosine diphosphate (ADP) and decreased ATP and NAD+/NADH in senescent cells.[31] Additionally, senescent cells do not proliferate but are resistant to autophagy and apoptosis and are thus long living. Importantly, senescent cells can exacerbate mitochondrial dysfunction, inflammation, and other disease-promoting pathways through SASP.[32] It has been debated whether senescent cells contribute to aging or are simply a protective mechanism against the development of inflammatory diseases. However, it is clear that senescent cells accumulated in tissues with aging may stop cell regeneration and tissue maintenance and are physiologically important drivers of age-associated functional decline, morbidity, and mortality.[33] Deletion of senescent cells from tissues of mouse models can delay or prevent multiple age-related diseases.[34] However, it remains elusive as to how senescent cells contribute to age-related diseases. SASP is emerging as a key driver of inflammation, and persistent inflammation may result in the accumulation of senescent cells in tissues, further contributing to age-related diseases.[35]
Autophagy/mitophagyAutophagy, a homeostatic process with multiple effects on immunity, has been recently recognized as a new hallmark of aging.[14] Autophagy is a mechanism in which the eukaryotic cell encapsulates damaged proteins or organelles for lysosomal degradation and recycling.[36] A growing body of evidence suggests that autophagic activity declines with age.[37] Furthermore, autophagy can modulate the major features of aging such as DNA repair and nutrient sensing/metabolism.[38] Autophagy can be initiated by calorie restriction, endoplasmic reticulum (ER) stress, or amino acid depletion through either the autophagy-related (ATG) or vacuolar protein sorting (VPS ) gene.[39] Mechanistically, studies demonstrated that Atg5 transgenic mice displayed anti-aging phenotypes that could extend the lifespan by enhancing autophagic activity.[40] Furthermore, Becn1F121A/F121A knock-in mice showed higher levels of basal autophagic flux and improved lifespan in both male and female mice.[41] In the meantime, human research targeted promoting autophagy to increase lifespan through lifestyle improvement and pharmacological modulation.[42]
Mitophagy is the selective degradation of mitochondria by autophagy. Mitophagy is an evolutionarily conserved homeostatic process by which the cells selectively degrade only dysfunctional or damaged mitochondria.[43] Mitophagy can act either as a response to nutrient starvation and oxidative stress or as a programmed removal of damaged mitochondria.[44] Mitophagy promotes the turnover of mitochondria and prevents the accumulation of dysfunctional mitochondria. Mitophaging was defined as the defective removal of damaged mitochondria through mitophagy resulting in degenerative diseases and aging.[45] There are substantial data on the effects of mitophagy changes on health and lifespan that support a decline in mitophagy and mitophaging in aging.[45]
Epigenetic modificationsEpigenetic alterations affect all cells and tissues throughout life. Loss of epigenetic information has been considered a cause of mammalian aging.[46] The epigenetic mechanism accompanied by cell senescence during aging has multiple modifications in DNA methylation, histone modification, chromatin remodeling, and non-protein-coding RNA transcripts (ncRNAs) that dictate their cell fate.[47] Endogenous DNA damage has been recognized as a significant causal factor in age-related systematic and stochastic changes in DNA methylation. DNA methylation can not only upregulate the expression of p21WAF1/Cip1 and p16INK4A22, but also induce DNA damage.[48] In addition to DNA methylation, epigenetic modifications of histones play an important role in DNA damage and aging. Histone modifications include alterations in phosphorylation, acetylation, and methylation, as well as chromatin remodeling, which change significantly with age. Notably, histone modifications have become a significant regulator and sensitive marker of telomere attrition, such as histone gamma H2A histone family member X (γH2AX).[49] The NAD+-dependent SIRT1, SIRT6, and SIRT7 as anti-aging medicines may catalyze modifications in histone proteins to regulate gene transcription and genome instability.[50] Recent advances have demonstrated that the senescence-associated ncRNAs can regulate cell proliferation and cell cycle arrest by affecting the transcripts of the inhibitor of CDK4/alternative reading frame (INK4/ARF)/very long integenic non-coding RNAs (vlincRNA) VAD/MIR31HG locus to increase p14ARF, p15INK4b, p16INK4a, and B-cell lymphoma-2 (Bcl-2).[51] Furthermore, some ncRNAs can also modulate the expression of SASP genes in senescent cells or directly interact with them. Different epigenetic modifications may interact with each other, coregulate gene expression, and eventually form an intricate network that is associated with aging.
Composition and diversity of the microbiotaCurrent researchers have focused on the role of the commensal human microbiome in health and disease. Over 10 microorganisms are inhabited in the human gastrointestinal tract and maintain the structure and function of a healthy gut ecosystem.[52] These microorganisms are essential for metabolic homeostasis, immune regulation, and protection against pathogens through different mechanisms, such as inducing interleukin (IL)-10 production, activation of Tregs (transforming growth factor-β [TGFβ]-producing regulatory T cells), T helper 17 (Th17) cells, and innate lymphoid cell type 2 (ILC2).[53,54] While the reason for this remains unclear, the composition of the gut microbiota of elderly humans markedly differs from that of young and middle-aged adults. For example, compared to the young population, the middle-aged adults showed decreased Actinobacteria and Firmicutes but increased Proteobacteria.[55] The compositional shift, together with age-associated loss of structural integrity of the gut, may contribute to the onset of immune dysregulation and manifestation of aging-associated pathologies.[56] In elderly people, the composition of the gut microbiota shows signs of dysbiosis. The decrease of beneficial microbes, particularly supporters of mucin production and producers of short chain fatty acids (SCFAs), appears to induce a chain of pathogenic inflammatory changes, thereby leading to inflammaging and aging-associated morbidities. Thus, gut dysbiosis and leakiness are major causes of increased mortality and premature death in elderly people. Recent advances in next generation sequencing technologies have allowed the identification of notable changes in the gut microbiota’s composition and diversity with age.[57]
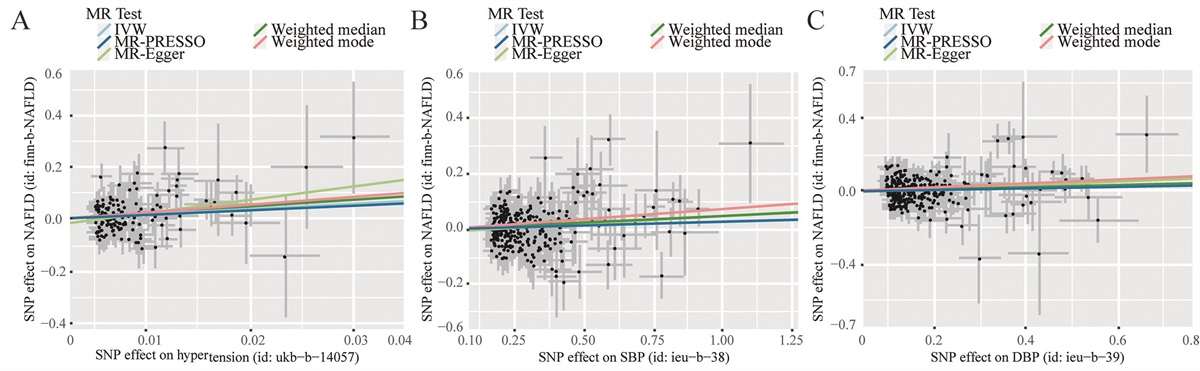
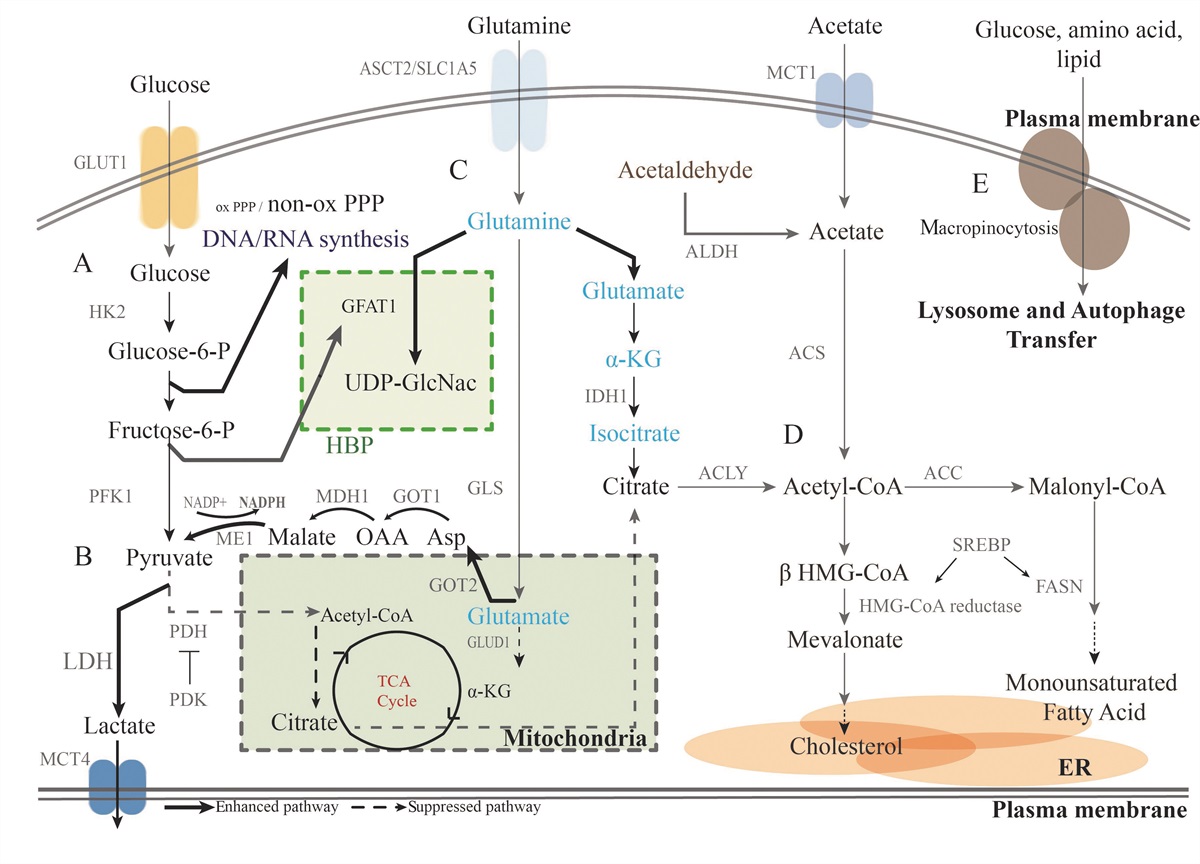
AhR and Hallmarks of AgingAhR has been considered initially as a receptor of polycyclic aromatic hydrocarbons (PAH) or dioxins; a lot of ligands have been recently identified in food or microbiota but are also produced by the human body (endogenous ligands).[58] Upon ligand binding, AhR translocates from the cytosol to the nucleus, leading to changes in target gene transcription (e.g., cytochrome P450 a1 [cyp1a1] and b1 [cyp1b1]) and immunotoxicological effects[59] [Figure 3]. AhR is featured not only in mediating the toxicity of dioxins, dioxin-like compounds (DLCs), and PAHs, but also in regulating a series of physiological functions.[60] Interestingly, the ligands of AhR from environmental (e.g., pollutants and other toxicants) and nutritional (e.g., flavonoids, carotenoids) factors influence aging and mitochondrial functions directly or indirectly.[61] Furthermore, expression of the AhR protein decreases with age, and lack of AhR has been associated with premature aging processes involved in increased cellular senescence and inflammaging features.[62] Additionally, we performed bioinformatical analyses on public datasets from the Gene Expression Omnibus (GEO) database GSE40732 with microarray data from peripheral blood mononuclear cells (PBMCs) of asthmatic patients and healthy controls. We found that AhR expression is highly correlated with several major hallmarks of aging, including epigenetic regulation, autophagy, immune response, DNA damage, protein destabilization, telomere maintenance, and defense against the bacterium [Figure 4]. Here, we review the existing literature on the role of AhR in regulating some of the hallmarks of aging: mitochondrial dysfunction, cellular senescence, autophagy/mitophagy, epigenetic regulation, and microbiome disturbance.
 Figure 3:
Figure 3: Model of the AhR signaling pathways. Upon binding to ligands from dioxins, allergens, bacterial-derived compounds, dietary sources, and pharmaceuticals, the AhR is activated and then translocated into nucleus. On one hand, AhR in the nucleus forms a heterodimeric complex with the ARNT and binds to a DRE consensus. This induces the expression of the AhR target genes, such as CYP1A1, CYP1A2, CYP1B1, AHRR, and COX-2, which are involved in a canonical signaling pathway. AhR can also be controlled via nuclear export and subsequent AhR degradation through the ubiquitin–proteasome signaling pathway. On the other hand, AhR in the nucleus can directly interact with other proteins such as the members of the NF-κB protein family or participate in the cross-talk with ER, which is involved in a non-canonical signaling pathway. AhR: Aryl hydrocarbon receptor; AHRR: Aryl hydrocarbon receptor repressor; ARNT: Aryl hydrocarbon receptor nuclear translocator; COX-2: Cyclooxygenase-2; CYP1A1/2: Cytochrome P450 1A1/2; CYP1B1: Cytochrome P450 1B1; DRE: Dioxin responsive elements; ER: Estrogen receptor; HSP90: Heat shock protein 90; NF-κB: Nuclear factor-κB; STAT: Signal transducer and activator of transcription; XAP2: Aryl hydrocarbon receptor interacting protein; XRE: Xenobiotic responsive element.
 Figure 4:
Figure 4: Association of AhR expression with hallmarks of aging. Bioinformatical analyses were performed on public datasets from GEO databases GSE40732 with microarray data from PBMCs of asthmatic patients and healthy controls. The samples are divided into AhR-High and AhR-Low groups based on the median expression value of AhR. GSEA analysis is performed with DEGs in a ranked manner. The NES are listed on the bottom of each bar. Only those biological processes with statistical significance (adjusted P-value <0.05) were illustrated, including epigenetic regulation, autophagy, immune response, DNA damage, protein destabilization, telomere maintenance, and defense to the bacterium. AhR: Aryl hydrocarbon receptor; DEGs: Differential expressed genes; GEO: Gene expression omnibus; GSEA: Gene set enrichment analysis; NES: Normalized enrichment scores; PBMCs: Peripheral blood mononuclear cells.
AhR and mitochondrial dysfunctionBoth environmental and nutritional factors can bind AhR and trigger the transcriptional activity of AhR, suggesting a possible link between AhR and mitochondrial dysfunction. Indeed, AhR signaling activated by 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) could cause oxidative stress and disrupt mitochondrial metabolism, leading to mitochondrial dysfunction. Similarly, benzo[α]pyrene can also promote mitochondrial dysfunction and inhibit the MMP, leading to the depletion of ATP levels and reduction of the oxygen consumption (OC) rate.[63] Our recent studies suggest that increased mitochondrial reactive oxygen species (mtROS) generation induced by allergens can promote mitochondrial dysfunction with decreased basal and maximal respiration and ATP turnover rate.[64] It is well recognized that elevated levels of mtROS may lead to mitochondrial dysfunction.[19] We have previously suggested that AhR signaling is critical in regulating mtROS generation that contributes to different inflammation-associated phenotypes, including mast cell degranulation and activation,[65] epithelial cytokine release,[66] and NLR family pyrin domain containing 3 protein (NLRP3) inflammasome formation.[67] Other studies also suggest a possible involvement of mitochondrial dysfunction and ROS generation in an AhR activation-dependent manner in a cellular model of non-alcoholic fatty liver progression induced by 6-benzylaminopurine/benzyl adenine (BaP)/ethanol co-exposure.[68] Mechanistically, particulate matter (PM) exposure can induce increased ROS production and reduced MMP with a lower expression of SIRT1, and the SIRT1 activator, SRT1720, can effectively inhibit ROS production and reverse PM-induced mitochondrial dysfunction.[69] SIRT1 and SIRT3 belong to NAD+ dependent deacetylases, are key to the control of metabolic processes and are localized to the nucleus and mitochondria, respectively.[70] Thus, this finding implicates that there might be a cross talk between AhR, Sirt1, and mitochondria that are essential for aging process.
AhR controls the balance of cellular senescence and reprogramingMitochondrial dysfunction has been associated with age-associated cell and tissue changes via different signaling pathways, thereby contributing to senescent phenotypes. Given that AhR signaling is critical in regulating mitochondrial function, it is possible that AhR also modulates cellular senescence. Indeed, kynurenine, as a tryptophan metabolite and an endogenous AhR agonist, has been reported to inhibit autophagy and promote senescence in aged bone marrow mesenchymal stem cells via AhR signaling.[71] This study suggests that AhR is a novel target to prevent or reduce age-associated bone loss and osteoporosis. In contrast, mouse embryo fibroblasts from AhR-/- mice showed earlier senescence than WT mice during adipogenic differentiation.[72] Further studies indicate that aged AhR-/- mice had exacerbated cellular senescence.[73] AhR signaling can also inhibit the proliferation of stem cells by repressing cell-cycle progression. Mechanistically, activated AhR can increase the expression of the p21Cip1 and p27Kip1 proteins and thus arrest cell proliferation. Moreover, AhR can bind elongation factor 2-promoter-binding factor (E2F) transcription factors and subsequently inhibit the transcription of many E2F-regulated genes, thereby controlling cellular senescence and aging. Collectively, the role of AhR in regulating senescence likely varies depending on different cellular and environmental contexts. Although both AhR and senescence have been shown to have beneficial effects in immune regulation and tolerance, they can also be detrimental when they lead to oxidative stress and DNA damage. Studies are clearly needed to uncover the diverse effects of AhR activation and its implications for cellular senescence and overall health.
AhR and autophagy/mitophagyThere is growing evidence that AhR could suppress autophagic activity in different cells through multiple mechanisms. For example, TCDD treatment can not only downregulate the expression of several autophagy-related genes in human keratinocytes, such as microtubule-associated protein 1A/1B light chain 3 (IC3), ATG5, and Beclin 1, but also suppress the formation of autophagosomes preventing autophagy-mediated cell death in the psoriasis pathogenesis.[74] In addition, the AhR agonist kynurenic acid (Kyn) has been shown to accumulate in plasma and tissues with age.[75] Kyn treatment can induce AhR nuclear translocation and disrupt autophagy, and inhibition of the AhR pathway prevented the kynurenine-induced increase of senescence and preserved autophagy in BMSCs in aged mice.[76] Of interest, the inhibition of autophagy can also increase the level of the AhR protein in different cell types (e.g., keratinocytes, HeLa cells). While the mechanisms remain unclear, recent studies suggest that glycogen synthase kinase 3β (GSK3β)-induced phosphorylation can promote the degradation of AhR protein via the autophagy-lysosomal pathway.[77] Several other studies have demonstrated that the ligand-induced activation of AhR factor stimulates the proteasome-dependent pathway, whereas the non-activated turnover of AhR protein is under physiological regulation via autophagy.[76] We have demonstrated a functional axis of AhR-ROS-NLRP3 inflammasome in regulating allergic airway inflammation.[67] The NLRP3 inflammasome has been associated with mitophagy-mediated maintenance of mitochondrial homeostasis. Thus, AhR signaling may be involved in regulating autophagy/mitophagy, which could be one of the possible mechanisms driving the aging process.
AhR and epigenetic regulationAhR can selectively bind to the unmethylated form of a specific sequence called the xenobiotic responsive element (XRE). The ligand-specific transcriptional response depends on AhR-DNA binding to XREs located in the regulatory regions of each gene. Studies have suggested that AhR is a novel DNA methylation reader, unlike classical methylation readers, such as methyl-CpG-binding protein 2, which binds to methylated sequences.[78] With exposure to endogenous AhR ligands, such as Kyn, methylation states of the individual target XREs are regulated to coordinate the expression of downstream genes that maintain homeostasis in a tissue-specific manner. However, continuous exposure to AhR ligands can cause changes in the methylation patterns around the XRE sequence and lead to different immune responses. Thus, it is likely that these environmental (e.g., pollutants and other toxicants) factors may induce DNA methylation through AhR-dependent mechanisms. In addition to DNA methylation, AhR impacts histone hyperacetylation and methylation through interactions with coactivators or by displacing histone deacetylase (HDAC) complexes. In turn, histone acetylation was found to be critical in the activation of the AhR promoter. Metastasis tumor-associated protein 2 (MTA2) was identified as a cofactor recruited by the AhR–aryl hydrocarbon receptor nuclear translocator (ARNT) complex exclusively in response to cinnabarinic acid (CA), an AhR agonist.[79] MTA2 is a chromatin-modifying protein and a component of the nucleosome remodeling and deacetylation complex with the capacity to repress and activate gene expression. A recent study demonstrated that CA-specific recruitment of the MTA2-AhR complex to XREs in the promoter of AhR target gene stanniocalcin 2 (Stc2) with concomitant acetylation of lysine 5 on histone H4 (H4K5Ac) at the Stc2 promoter leads to the transcriptional activation of target genes.[80] Additionally, activation of AhR is tissue-specific and depends on chromatin-accessible regions. These findings indicate that AhR is a pivotal player in epigenetic regulation in response to age-associated environmental and nutritional factors.
AhR as a mediator of host–microbiota interplayThe cumulated evidence demonstrated that the activation of AhR could be either beneficial or detrimental to aging.[81] For example, AhR can be predominantly activated by ligands produced from gut microbes metabolizing diet-derived tryptophan. The gut microbiota can metabolize tryptophan to activate AhR signaling, which participates in varying physiological processes rather than pathophysiological events associated with aging.[82] However, aberrant tryptophan metabolism and dysbiosis of gut microbiota may lead to changes in AhR activity during aging that might contribute to the acceleration of the aging processes.[83] Similar to the “imbalance” in the gut microbial community, dysbiosis of the microbiota in the gut, “dysbiosis” of the skin microbiota is equally important by affecting skin homeostasis, exacerbating a variety of skin diseases, and affecting disease diagnosis and treatment. For example, Staphylococcus aureus (SA) was found on the skin of patients with atopic dermatitis (AD) at the site of lesions caused by AD that can impede the function of the skin barrier or activate the inflammatory response.[84] A recent study demonstrated that SA could activate the AhR–ARNT system and enhance the terminal differentiation of epidermal keratinocytes.[85] However, the metabolites that served as AhR ligands remain unidentified. Further, AhR activation could upregulate the expression of antimicrobial peptides in keratinocytes, contribute to the changes in skin microbiota composition, and restore dysbiosis in AD.[86] Thus, AhR and tryptophan metabolites are important regulators in the “microbiota–AhR–skin” axis.
AhR signaling, Hallmarks of Aging, and AsthmaAsthma is recognized as a comorbidity of aging because aging can influence the occurrence and development of asthma.[87] Intriguingly, the hallmarks of aging, such as mitochondrial dysfunction, telomere shortening, epigenetic alterations, and altered intercellular communication, have been observed in patients with asthma.[88] We also performed bioinformatical analyses on the gene expression omnibus (GEO) database GSE69683 for the relationships between asthma and the hallmarks of aging [Figure 5]. Our analyses provided supporting evidence that asthma is associated with these hallmarks of aging. Here we provide an overview of the impact of aging hallmarks on asthma phenotypes (e.g., airway inflammation, remodeling, airway hyper-responsiveness, asthma exacerbation, and therapeutic response).
 Figure 5:
Figure 5: Association of asthma with hallmarks of aging. Bioinformatical analyses were performed on public datasets from GEO databases GSE69683 with microarray data from PBMCs of asthmatic patients and healthy controls. The comparison is performed between asthma patients and relative normal control. GSEA is performed with DEGs in a ranked manner. The NES are listed on the bottom of each bar. Only those biological processes with statistical significance (adjusted P-value <0.05) were illustrated, including immune response, DNA damage, epigenetic modification, mitochondria, protein stabilization, and autophagy in asthmatic patients. DEGs: Differential expressed genes; GEO: Gene expression omnibus; GSEA: Gene set enrichment analysis; mRNA: Messenger RNA; NADH: Nicotinamide adenine dinucleotide hydrogen; NES: Normalized enrichment scores; PBMCs: Peripheral blood mononuclear cells.
“Elderly” was defined as the chronological age of 65 years or more. Elderly patients with asthma not only had a higher prevalence but also had the highest death rate at 5.8 deaths per 10,000 people.[89] For those elderly patients, there are significant changes in the innate and adaptive immune responses to environmental exposures that contribute to an increas
Comments (0)