
Remember me




There were 3899 individuals across 7 cohorts available for maternal asthma during pregnancy and 7433 individuals across 13 cohorts for maternal asthma ever. Supplementary material contains cohort-specific asthma definitions and distributions of parental asthma phenotypes. The prevalence of self-reported maternal asthma ranged from 2 to 35%, sustained smoking ranged between 0.4 and 30%. Most cohorts used the 450 K chip. Two cohorts (NorthPop and UpstateKIDS) had DNA methylation measured using the EPIC chip. Participants were primarily of European descent (Table 1). We did not see any overlap between our findings and a recently published EWAS on maternal BMI [22].
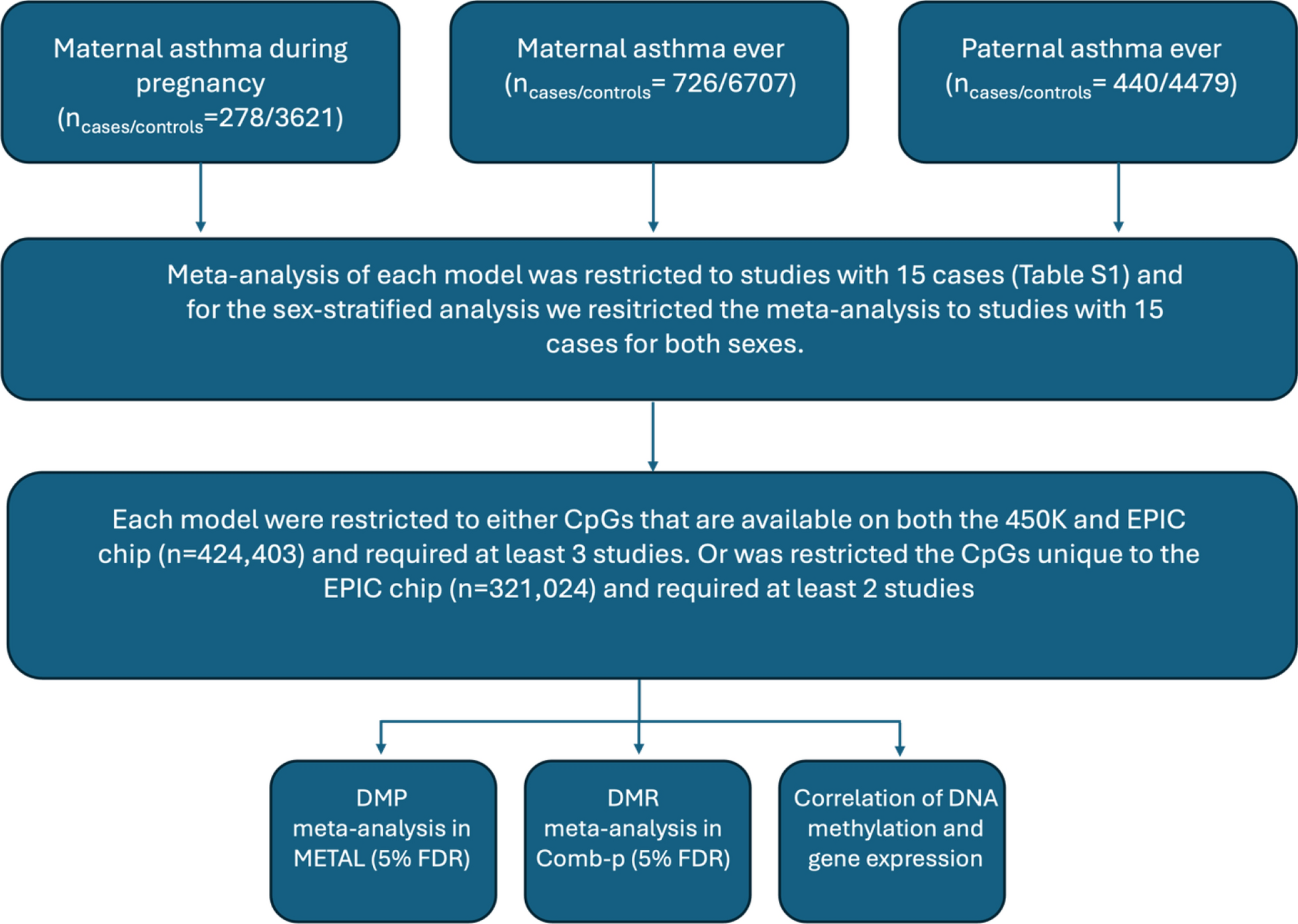
Table 1 Characteristics of participating study cohortsAn overview of included analyses, primary quality control filters and analyses are shown in Fig. 1.
Fig. 1
Flowchart describing the number of samples, the primary quality control filters as well as the analyses included
Maternal asthma during pregnancy and newborn DNA methylationThe meta-analysis of newborn DNA methylation in relation to maternal asthma during pregnancy included 278 exposed and 3621 non-exposed participants from 7 cohorts: IOW, EDEN, MoBA1, MoBA2, NorthPop, UpstateKIDS and MINERvA. We identified 1 significant DMC for the 450 K chip (λ = 1.09), cg26963854 within the south shelf of a CpG island, which was not annotated to a specific gene (FDR 5%). We also identified 26 DMCs exclusive to the EPIC chip (FDR < 5%) using 76 exposed and 675 non-exposed participants from NorthPop and UpstateKIDS (λ = 1.18) (Fig. 2, Table 2). Using a stricter P value threshold of (FDR < 2.5%), we observe 6 DMCs pertaining only to the EPIC chip (Table S2). We identified 5 DMRs (encompassing 28 CpGs) in relation to maternal asthma during pregnancy for the 450 K chip and none for probes exclusive to the EPIC chip Table S3). Forest plots with cohort-specific beta values and 95% confidence intervals for the identified CpGs are shown in Fig. S1. For the CpG site (cg26963854) identified using the 450 K chip, the effect estimate was lower in the MINERvA cohort compared to that of the other included cohorts with the 450 K data, but without evidence of heterogeneity (Pheterogeneity = 0.14). For the EPIC chip, we saw no evidence of heterogeneity (Fig. S1).
Fig. 2
Manhattan plot of epigenome-wide association of maternal asthma during pregnancy and offspring methylation. Models include the following covariates: child sex, maternal smoking during pregnancy, gestational age, mode of delivery, maternal age at childbirth, maternal socioeconomic status, estimated cell type, batch covariates and ancestry. Panel A) shows analysis restricted to 435,329 probes on the 450 K chip for 7 cohorts using 202 exposed and 2946 non-exposed individuals (lambda = 1.09), and panel B) shows 321,034 probes exclusive to the EPIC chip for 2 cohorts using 76 exposed and 675 non-exposed individuals (lambda = 1.18). The red line indicates 5% FDR significance
Table 2 Twenty-seven differentially methylated CpGs (FDR < 0.05) from the meta-analysis of maternal asthma during pregnancy in relation to newborn methylationMaternal asthma ever and newborn DNA methylationThe meta-analysis of newborn methylation and maternal asthma ever included 726 exposed and 6707 non-exposed participants from 13 cohorts: ALSPAC, CHS, DCHS, EDEN, GENR, INMA, IOW, LiNA, MINERvA, MoBA1, MoBA2, NorthPop and UpstateKIDS, and identified no associated probes (FDR < 0.05) for either the 450 K or EPIC chip (Fig. S2. We identified 3 DMRs (comprising 28 CpGs) for the 450 K chip but none for the EPIC chip (Table S4). We did not observe any significant probes (FDR < 0.05) if we restrict the maternal asthma ever analysis to the 7 cohorts in the maternal asthma during pregnancy analysis.
Paternal asthma and newborn DNA methylationThe meta-analysis of newborn methylation and paternal asthma ever included 440 exposed and 4479 non-exposed participants from 9 cohorts: ALSPAC, GENR, INMA, IOW, LiNA, MoBA1, MoBA2, NorthPop and UpstateKids, and identified no probes associated with paternal asthma (FDR < 0.05) for the 450 K chip and 2 associated probes for the EPIC chip, namely cg08311378 in the gene body of RPS6KA2 and cg07462855 in the gene body of FAM160B1 (Fig. S3, Table S5).These DMCs were not among the 26 DMCs identified using the EPIC chip for the analysis of maternal asthma during pregnancy nor were they at least nominally significant in the maternal asthma (active or ever) analysis. Forest plots for the identified DMCs are in Fig. S4.
We identified 11 DMRs encompassing 99 CpGs in relation to paternal asthma ever diagnosis for the 450 K and 2 for the EPIC chip, encompassing 9 CpGs (Tables S6). Some overlap was detected with DMRs associated with maternal pregnancy asthma status (in gene PPT2; PRRT1) and maternal ever asthma (HOXA genes).
Correlation analysis of newborn methylation across maternal and paternal asthmaWe correlated methylation effect sizes across 450 K CpGs from the meta-analyses results from maternal asthma during pregnancy (MAP), maternal asthma ever (MAE), maternal asthma ever without using MAP data (MAE exclusive) as well as paternal asthma ever (PAE) (Fig. 3). We observed a moderately strong positive correlation between MAP and MAE effect sizes (rho = 0.55, P value < 0.001), as well as positive correlation between MAP and MAE exclusive effect sizes (rho = 0.47, P value < 0.001). We did not observe a correlation between MAP and PAE (rho = 0.002, P value = 0.15), but observed a weak positive correlation between MAE and PAE (rho = 0.05, P value < 0.001) and p between MAE exclusive and PAE (rho = 0.06, P value < 0.001).
Fig. 3
Spearman correlations of effects from meta-analyses of all included models for A) CpGs pertaining to the overlap between the 450 K and EPIC chip and B) the CpGs unique to the EPIC chip. MAP, maternal asthma during pregnancy; MAE, maternal asthma ever while; PAE, paternal asthma ever. Sex-stratified models are defined by suffixes. ‘Int’ represent interaction models testing for differences between the sexes
For the EPIC chip, MAP and MAE effect sizes we again found moderate positive correlated (rho = 0.47, P value < 0.001). MAP and PAE were weakly negatively correlated (rho = − 0.04, P value < 0.001) as were MAE and PAE (rho = − 0.03, P value < 0.001) (Fig. 3).
The single DMC found on the 450 K chip, cg26963854 located on chromosome 14, that passed the FDR threshold of 5% in maternal asthma during pregnancy had a similar direction of effect across the maternal asthma ever and paternal asthma ever models but was only nominally significantly associated (P < 0.05) in the maternal asthma ever meta-analysis.
Of the 26 DMCs identified in maternal asthma during pregnancy meta-analysis on the EPIC chip, we focused on the 21 that were available across the maternal and paternal asthma ever models. We observed similar direction of effect in the maternal asthma during pregnancy and the maternal asthma ever meta-analysis for 19 of these 21 CpGs, and among these 19, we observed 13 nominally significant p values (Table S7). In contrast, none of the CpGs identified in the maternal asthma during pregnancy analysis had significant p values in the paternal asthma ever model nor consistent directions of effect (Table S7). Of the two DMCs, cg08311378 and cg07462855, identified in paternal asthma ever meta-analysis on the EPIC chip, we did not observe a similar direction of effect when compared to effect sizes in MAP (Table S8). We also did not observe nominally significant p values in the MAP meta-analysis results. Compared to the MAE analysis effect estimates, we saw that cg07462855 had a similar direction of effect but neither of the DMCs were nominally significant (Table S8).
CpGs associated in the literature with childhood asthma or pulmonary functionWe uploaded the top CpGs on the 450 K chip identified for MAP (Table S9), MAE (Table S10) and PAE (Table S11) to the EWAS toolkit platform [30] to investigate enrichment in previous DNAm results. For the trait enrichment analyses using MAP-associated CpGs, we observed enrichment in the following traits (which were also among the top10 most associated traits): asthma, smoking, atopy and maternal smoking (all had enrichment P values < 1.98 × 10–13) (Table S12).
Similarly, we observed that asthma and smoking were the top two traits among all traits when using MAE-associated CpGs (Table S13). Using CpGs associated with paternal asthma ever, we also observed significant association with asthma and smoking (Table S14).
In addition, we investigated enrichment for DMCs identified in the literature specifically for childhood asthma and lung function [8, 11, 31, 32]. We included DMCs identified in blood in an investigation of neonates developing asthma and among children with a clinical diagnosis of asthma in Reese et al. 2019 [8], DMCs in whole blood from childhood asthma in Xu et al. 2018 [31], DMCs identified in nasal epithelial cells from Qi et al. 2020 [32] and unique DMCs identified across DMRs for FEV1, FEV1/FVC and FEF75 in cord blood [11]. In total, 766 previously identified DMCs were used as the enrichment target, and we considered significant enrichment using a P value cutoff of 5% from Fisher’s exact test and used as input the CpGs in our analyses with P values below 0.005 (Table S9, S10 and S11). We did not observe any enrichment across maternal asthma during pregnancy (P value = 0.57), maternal asthma ever (P value = 0.84) or paternal asthma ever (P value = 0.85).
Sex-specific analysesSex-stratified analyses were performed for all 3 main phenotypes. For the sex-stratified meta-analysis of newborn methylation and maternal asthma during pregnancy, we included 102/943 exposed/non-exposed for boys and 83/781 exposed/non-exposed for girls from 3 cohorts: MoBA1, MoBA2 and NorthPop. We observed 6 DMCs for boys and 325 DMCs for girls related to maternal asthma during pregnancy at FDR < 0.05. None of the 6 DMCs identified in boys gave evidence of significant interaction with sex (FDR < 0.05) (Table S15. Investigating the 325 DMCs identified for girls, 154 had nominal significance for the sex-specific interaction; however, none was statistically significant (FDR < 0.05). Of these 325, 19 were also nominally significant (14 had same direction of effect) in the boys and not among the 6 DMCs identified in boys alone (Table S16).
For the sex-stratified meta-analysis of newborn methylation and maternal asthma ever, we included 309/2347 exposed/non-exposed for boys and 278/2206 exposed/non-exposed for girls from 7 cohorts: ALSPAC, GENR, LiNA, MoBA1, MoBA2, NorthPop and UpstateKIDS; we identified 1 DMC in boys and 25 in girls; none gave evidence of interaction with sex (FDR < 0.05) but 15 of the DMCs identified for girls met nominal significance (Table S17). For paternal asthma exposure, we included 183/1920 exposed/non-exposed for boys and 182/1834 exposed/non-exposed for girls from 7 cohorts: ALSPAC, GENR, LiNA, MoBA1, MoBA2, NorthPop and UpstateKIDS, and we observed 100 DMCs in boys and 95 DMCs in girls (Table S18, S19). Among the 100 DMCs identified in boys only, we saw nominally significant evidence of interaction for 5 sites (Table S18). Among the 95 DMCs found in girls, we observed ten DMCs, three with known annotation near genes HCCA2, C1orf198 and PNMT, with statistically significant interaction (FDR < 0.05) (Table S19). For meta-analysis of sex interaction for paternal asthma exposure, we identified 12 DMCs (Table S20), where we observed in general lower methylation and a stronger effect in girls compared to boys.
Differential DNA methylation and gene expression in blood and lungTo investigate whether differently methylated sites may be associated with gene expression, we analyzed eQTM pairs for 832 blood samples available from the HELIX consortium for child blood [29] and eQTM pairs for 424 lung samples from the GTEx consortium [33].
Among the 402 DMCs identified across all models using the 450 K chip, we observed 15 unique DMCs with significant associations with gene expression in blood (Table S21). Among these associations, we observed a decreased expression of FAM43A with increased methylation of cg02072170, and this gene is associated with eosinophil counts and thus associated with asthma etiology [34]. We also observed increased expression of LTBP1 with increased cg15772133 methylation, and this gene has been associated with FEV1/FVC in adults [35, 36].
Among the 441 unique DMCs that are encompassed by identified DMRs across all models using the 450 K chip, we found 188 significant unique methylation and gene expression pairs using the HELIX data (Table S22). We observed CpGs that annotated to genes that previously have been associated with asthma in adults including HLA genes [37].
Examining 56 DMCs identified from DMRs in models with maternal asthma during pregnancy and ever as the primary exposures, we found 35 unique DMCs were associated with gene expression of 4 genes. One gene of particular interest is HOXA5, which has previously been associated with organogenesis [38], lung function in adults [39, 40] and mental disorder phenotypes [41] (Table S22). The two other identified genes (KDM2B and KCTD11) were found to be involved with neurodevelopmental disorders [42] and cancer [43], respectively.
Among the 402 unique DMCs identified across all models using the 450 K chip, we observed one association between increased methylation of cg20810675 and decreased expression of C4orf27 in lung tissue in GTEx (Table S23). Among the 411 DMCs that are encompassed in DMRs across all models, we identified substantially more associations with gene expression compared to the 402 unique single DMCs identified (Table S24). Specifically, these associations included many HLA gene variants and also the NOTCH4 gene which has been associated with schizophrenia [44], psoriasis [45] and asthma [46].
Druggable targetsWe identified differential methylation in regions related to parental asthma involving the HOXA5 and HLA genes. HOXA5 is also the target of CHEMBL4224852, a lysine demethylase, which may implicate epigenetic regulation in asthma development. Several HLA genes were also identified as targets for drugs, including HLA-C (target of CHEMBL4680046) and HLA-DRB1 (target of CHEMBL2109447).
Comments (0)