
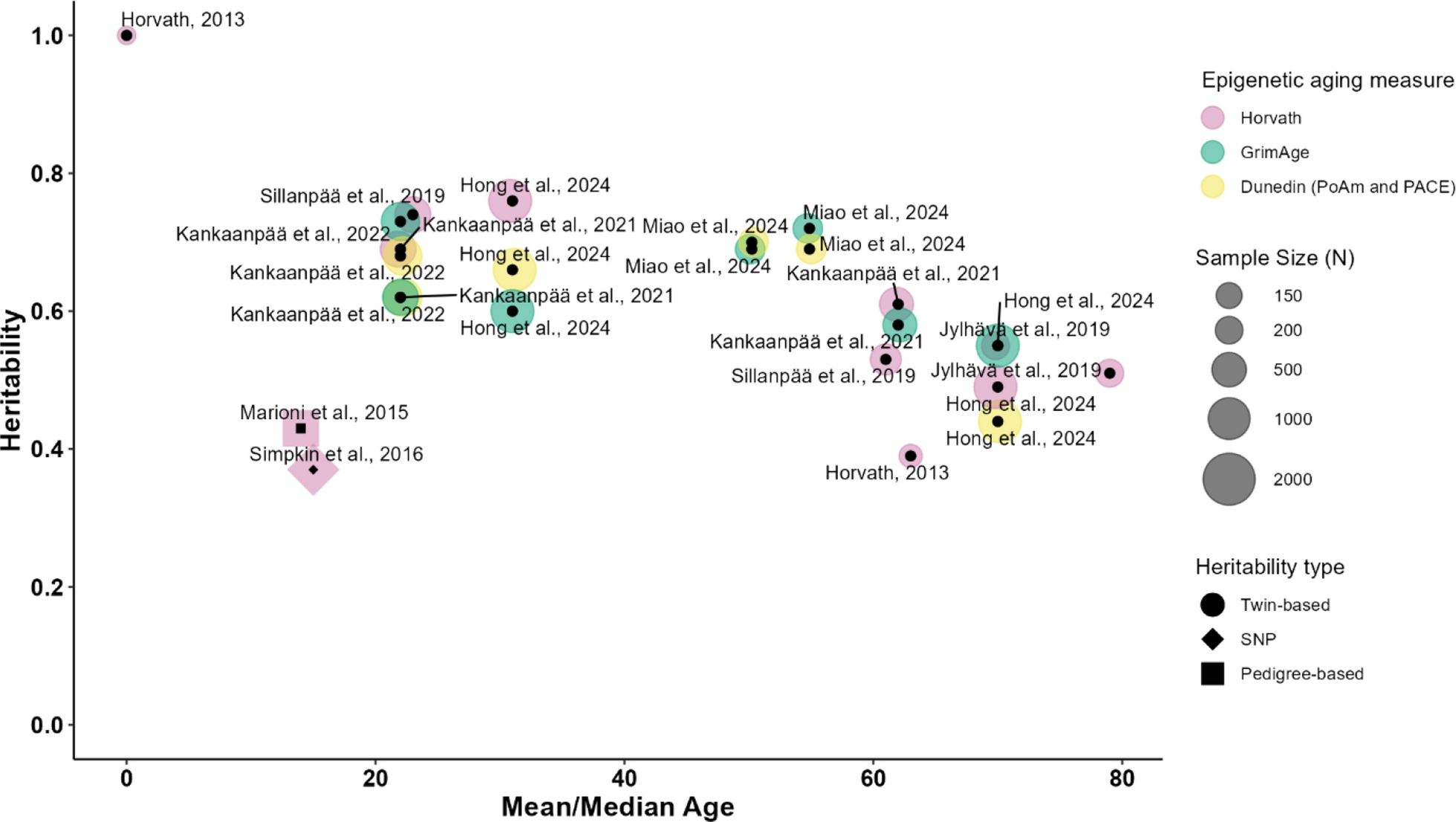
Remember me




In this study, cochleae were collected from six-week-old (6 W) and twelve-month-old (12 M) C57BL/6J mice. The mice evaluated in the experiment received ABR measurements at varying frequencies (4, 8, 16, 24, 32 kHz) and click to determine the level of hearing loss. As shown in Fig. 1A, hearing thresholds were significantly higher (two-way ANOVA, p < 0.0001 or 0.001) in 12 M mice (n = 17) than in 6 W mice (n = 12) at all frequencies and clicks. Interestingly, there were large differences among individual 12 M mice at varying auditory frequencies, consistent with what is observed in humans [5]. Earlier studies have indicated that C57BL/6 J mice display the characteristic pattern of ARHL between the ages of 12 and 15 months, which aligns with our findings mentioned above [28]. We therefore selected the 12 M mice (aging group) and 6 W mice (control group) for subsequent experiments.
Fig. 1
Histological characterization changes in aging cochleae and the identification of an age-related hearing loss mouse model. A ABR thresholds were observed in 6 W (n = 12) and 12 M (n = 17) C57BL/6 J mice at 4, 8, 16, 24, 32 kHz and click. B Young adult and old aged C57BL/6 J cochlear representative cross sections stained with H&E showed atrophy of SV with age in all three turns. Section thickness is 5 μm. C The quantified analysis of SV in three turns, which showed the most SV atrophy appear in basal turns. D, E, F The difference in HCs counts of young adult and old mice at same locations was calculated, and dramatic loss of HCs in cochlear basal turn was observed. (n = 3). G, H Apoptotic loss of SGCs was detected in two groups and higher TUNEL fluorescence (green color) ratio of SGCs area was detected in 12 M mice than in 6 W mice between the same turn. Blue shows DAPI staining of the nucleus. Scale bar = 50 μm. I, J Western blot analyses of laminb1 and H3K4me3 in the cochleae from 6 W (n = 6) and12 M (n = 6) mice. The statistical significance was represented as * p < 0.05; **p < 0.01; ***p < 0.001;****p < 0.0001. Bar graph results are means ± SD from 3 independent experiments
Several experiments were performed to identify potential causes of ARHL and confirm that the observed hearing loss was indeed senescence-related. H&E staining of cochlear cross sections revealed age-related atrophy of the SV in all three turns (anatomy shown in Fig. 1B, cross section), as previously noted in prior investigations [29, 30]. Notably, cochlear cross sections showed that among the three turns, the most obvious SV atrophy appeared in basal turns (Fig. 1C), which is consistent with the characteristic high-frequency hearing loss in ARHL. HCs are sensory cells located within the cochlea of the inner ear, the loss of which is a common cause of hearing loss. The dissected cochlear sensory epithelium was stained with anti-myosin 7a antibodies to assess aging-associated HC loss. Obvious loss of IHCs was observed in apical and basal turns (Fig. 1D). A dramatic loss of OHCs in all three turn was observed, especially the basal turn (Fig. 1D). The quantitative analysis showed that HC counts were decreased in the aging group compared with the control group (Fig. 1E, F, n = 3). Progressive apoptotic loss of SGCs in the aged mice was detected by TUNEL staining (Fig. 1G). The TUNEL fluorescence ratio of the SGC area in the same turns showed statistically significant differences between 6 W and 12 M mice (Fig. 1H). TUNEL-positive SGCs were observed in aging mice, whereas most SGCs of control mice were TUNEL-negative. Moreover, a significant increase in the number of apoptotic SGCs in the basal SGC area compared with the middle SGC area was observed in aging mice, differing from the condition in 6 W mice.
Given the lack of specificity of senescence markers, a combination of different markers is always expected [31]. Downregulated levels of LaminB1 (a structural protein of the nuclear lamina) and H3K4me3 (associated with active transcription) have become common markers of senescence [32, 33]. Western blot analysis of cochleae showed that the LaminB1 and H3K4me3 expression levels of the 12 M mice were significantly lower (n = 12, Student’s t test, both p < 0.0001) than those of the 6 W mice (Fig. 1I, J). Based on the above findings, we have fully specified a successful ARHL mouse model.
Genome-wide chromatin accessibility profiling of the cochleae in ARHL and normal miceChromatin accessibility refers to the degree of compactness or openness of the chromatin structure, which can influence the accessibility of regulatory elements, such as enhancers, to transcription factors and other regulatory proteins. To ascertain chromatin-level epigenetic transcriptional regulation, we performed ATAC-seq to construct genome-wide maps of ACRs for the two groups (control group and aging group). Three biological replicates were performed for each group to ensure the consistency and reliability of the results. And the fraction of reads in peaks (FRiP) score was provided for ATAC-seq library quality (Table S2). The similarity within each group is shown by heatmap clustering of Pearson correlation coefficients from the comparison of 6(> 20FRiP)ATAC-seq profiles (Supplementary Fig. S1A). The within-group correlation in the aging and control groups was greater than the between-group correlation, suggesting that the ATAC-seq data are reliable.
An average of 131.16 million raw reads was obtained from all samples, and more than 98% of these were clean reads (Table S3). The GC content of clean reads in each sample was higher than 40% and showed a normal distribution (Fig. S1B). There were obvious enrichment peaks around the transcription start sites (TSSs), and the average sequencing depth in the TSS of the control group was higher than that of the aging group (Fig. 2A). To detect changes in chromatin accessibility during the cochlear aging process, we performed a differential analysis of the peaks. In total,1,309,220 and 1,249,876 peaks (or chromatin-accessible regions) were identified in control samples and ARHL samples, respectively (Figure S1D), and genome-wide chromatin accessibility of each is shown in Fig. 2B and Figure S1C.93779 peaks were identified as differential peaks (Table S4), which consist of 34,269 increased and 59,511 decreased peaks (Fig. 2C). The distribution plot of peak length revealed that the fragments detected in aging cochleae were shorter overall than those detected in control cochleae (Fig. 2D). These results implied that the overall chromatin accessibility decreased in the aging group during the aging process at the chromosomal level. Genomic annotation of the peaks showed that 2.74% of peaks were located in the promoter regions of genes (± 3 kb of TSSs), 36.18% were in introns, 58.32% were in intergenic regions, and 2.25% were in exons (Fig. 2E). GO and KEGG analyses revealed that the upregulated genes associated with differential peaks between the two groups mainly functioned in the cytoplasm and axons, participating in ‘immune response,’ ‘innate immune deficiency,’‘intestinal immune network IgA production,’ and ‘NF-κB signaling pathway’ (Fig. S2A-B, Table S5-6). GO and KEGG analyses of downregulated genes associated with differential peaks showed that they mainly functioned in the extracellular space and axoplasm, associated with ‘cilium movement,’ ‘innate immune deficiency,’ ‘NF-κB signaling pathway,’ ‘PI3K-Akt signaling pathway,’ and ‘mineral absorption’(Fig. S2C-D, Table S7-8). The x-axis represents the significance of enrichment (represented by -log10pvalue, where a higher value indicates more significant enrichment), while the y-axis represents the enriched terms. Here, we plotted the top 20 most significant terms according to ascending p-values.
Fig. 2
Genome-wide accessible chromatin profiling of cochleae of ARHL and normal mice. A Enrichment peaks around the transcription start sites (TSSs), and comparison of the average sequencing depth in TSS of two groups. (X: the distance from the site to the TSS, Y: the average sequencing depth of the site). B Genome-wide chromatin accessibility of control and aging cochlea. C Volcano plot of differential analysis to the peaks during the aging process of cochlea, which consist of 34,269 increased and 59,511 decreased peaks. D The distribution plot of the fragments length revealed that the fragments detected in aging cochleae were shorter overall than in control cochleae. E Genomic annotation of identified different ATAC-seq peaks. F The enrichment heatmap of nucleosome binding sites revealed a looser or more relaxed chromatin structure in the control cochleae. G The geneplot of Hmgb1 in different bioreplicated samples (the arrow indicates the transcription start site and direction, the green region represents the exonic region, and the higher the peak, the stronger the accessibility of that region)
Chromatin accessibility is determined by the local nucleosome occupancy and binding of chromatin-related proteins. Active cis-regulatory elements, such as promoters and enhancers, are located in open genomic regions, also known as nucleosome-depleted regions (NDRs). These regions are sensitive to nuclease activity, and thus are also termed DNase I hypersensitive sites, and can be identified by ATAC-seq. In this research, by integrating the structural characteristics of nucleosomes and the distribution of ATAC-seq inserted fragments, fragments within 105 bp are designated as NDRs, and inserted fragments of 105–250 bp are designated as nucleosome distribution areas, allowing precise nucleosome positioning. The heatmap presents the enrichment of nucleosome binding sites on either side of the core binding region of transcription factors in the control and aging cochleae (Fig. 2F). The read density of fragments within 105 bp in the control group was higher than that in the aging group, revealing a looser or more relaxed chromatin structure in the control cochleae, with active gene regulation and transcriptional activity. Additionally, the gene plots of a few well-known senescence markers, including LaminB1, Hmgb1, Egfr, and Mmp2, showed significantly different peaks in their promoter regions (Fig. 2G, Fig. S3A).
Motif enrichment analysis and motif-TF-gene prediction associated with ARHL progressionChromatin accessibility plays a key role in determining the binding of transcription factors to regulatory sequences [34]. By analyzing the binding motifs that are enriched within the regulatory regions of genes implicated in ARHL, we can identify potential TFs that may play crucial roles in the disease. Once enriched motifs are identified, motif-TF-gene prediction can be performed to associate TFs with specific ARHL-associated genes.
According to the DNA sequences of different peaks, the top 10 most significantly enriched TF binding motifs were known binding sites for CTCF, SP1, TBR1, ELF, KLF6, BORIS, SP5, KLF10, NFIL3, and KLF1 (Fig. 3A). The significant enrichment of these motifs suggests their importance in regulating gene expression in the context of ARHL. Indeed, during ATAC-seq library construction, DNA sequences that are directly occupied by DNA-binding proteins can be protected and thus less accessible to the transposase enzyme that fragments and tags accessible chromatin regions. This protection leads to the elimination or reduction of ATAC-seq signals within these regions, resulting in a characteristic sequence "footprint" or decreased read density. To further characterize the predicted key TFs involved in ARHL development, we performed a TF footprinting analysis for each TF, which enabled us to identify potential genome-wide TF binding events. The results of the analysis demonstrated a noticeable change in footprints across the 10 TFs studied (Fig. 3B). Specifically, the footprints of all 10 TFs displayed a deeper, more pronounced pattern in the control group than in the aging group, revealing that there were alterations in the protein‒DNA interactions and accessibility of the chromatin regions between the two groups. The increased footprint size or insertion-site probability indicates a higher likelihood of transposase enzyme insertion events occurring in the control group compared to the aging group. This suggests that there is a higher degree of protection for DNA sequences bound by TFs in the control group, resulting in increased accessibility and increased transposition during ATAC-seq library construction.
Fig. 3
Motif enrichment analysis and motif-TF-gene prediction associated with ARHL progression. A The top 10 significantly enriched TF binding motifs by decreased peaks from young to aging according to the enrichment p-values. The corresponding binding motif of TFs is shown. B Aggregated footprint plots display the top 10 TFs with reduced footprints, showing an overall footprint for all related transcription factor binding sites for each TF, with individual plots centered on binding motifs. C TF-gene regulatory network involved in ARHL progression extracted from HOMER data based on TOP 10 motifs. D The top 10 enriched GO terms by the genes regulated by predicted TFs indicated that the predicted TF-regulated genes were significantly enriched in different developmental processes
In order to study possible TF binding events in different periods, motif analysis was carried out on identified difference accessibility peaks, and matching binding TFs were extracted from HOMER data based on TOP 10 motifs, thus obtaining the regulatory network diagram of TFs and genes associated with difference peaks involved in ARHL progression (Fig. 3C, Table S9). Motif-TF-gene prediction analysis can help uncover potential TFs that may interact with these motifs and regulate the expression of ARHL-related genes. Each TF is represented as a red node in the network, and the edges between the nodes represent potential regulatory relationships between the TFs and target genes. The GO-KEGG pathway enrichment analysis of the TF network indicated that the predicted TF-regulated genes were significantly enriched in different developmental processes (such as urogenital system, renal system, and cardiac muscle tissue development and regulation of cell growth), regulation of kinase activity, DNA-binding transcription repressor activity, and negative regulation of the Wnt signaling pathway (Fig. 3D). This suggests that the TFs in the network may be involved in controlling key signaling pathways and transcriptional regulatory mechanisms through the regulation of kinase activity and repressing gene transcription.
Genome-wide gene transcriptional analysis comparing control and aging cochleae and prediction of ARHL-related TFsTo identify global gene expression changes between the two groups, we performed RNA sequencing using the cochlear tissues of three pairs of control and aging mice. An average of 35.75 million clean reads (accounting for 84.04%, Table S10) and a 98.54% mapping ratio were obtained from our deep RNA-seq libraries (Table S11). Hierarchical clustering based on gene expression levels divided the samples into two distinct clusters (red: upregulated, blue: downregulated) corresponding to the control and aging mice (Fig. 4A). In addition, we provide a higher-resolution display of the genes with significant differences between the two clusters (including gene names) in the original image for a more intuitive, in-depth analysis of the gene expression patterns and key references (Fig. S4). Differential analysis identified 155 and 399 significantly upregulated and downregulated genes, respectively (Fig. 4B, Table S12). These data reveal transcriptome changes during the development of ARHL. GSEA of 18,381 genes showed that 8301 (45.2%) genes with a correlation area of 38.5% were more highly expressed in aging cochleae, and 10,080 (54.8%) genes with a correlation area of 61.5% were more highly expressed in control cochleae. Then, we found that 1731 gene sets were upregulated in the aging group and 3442 gene sets were upregulated in the control group. GSEA enrichment results identified regulation of the B-cell receptor signaling pathway, regulation of the antigen receptor-mediated signaling pathway, axoneme assembly, appendage development, and cilium movement as gene sets with high enrichment scores (Fig. 4C). Among these gene sets, the first two were significantly enriched in the aging group, while the last three were significantly enriched in the control group. In addition, we found that the gene sets of various developmental processes, WNT–β-catenin signaling, ciliary plasm, and TNFA signaling via NFKB were also significantly enriched, which corresponded to the GO enrichment results of ATAC-seq. To explore the potential TFs related to the cochlear aging process, we performed TF prediction using the transcriptomic data (Table S13). Among all the predicted TFs, 9 were from the homeobox family, 3 were from the Fz-C2H2 family, and 3 were from the ETS family (Fig. 4D). The analysis of footprint changes of the top 5 TFs (|log fold change|> 1.8), including Phox2b, Pitx1, Pax1, Foxa1, and Meox1, indicated that the footprints of the 5 TFs were significantly decreased from young to aged mice genome-wide, which was in accord with the TF predictions from the transcriptomic data (Fig. 4E).
Fig. 4
Genome-wide gene transcriptional difference analysis between the young and aging cochleae and TFs prediction related to ARHL. A Heatmap analysis and hierarchical clustering on transcriptome samples using normalized read counts (TPM values). Each row represents a gene, and the TPM values were Z-scaled by row. The scale bar indicates the Z-scaled TPM values. B The scatter plot of differential expression genes. Gray dots represent genes that are not differentially expressed, blue dots represent genes that are differentially downregulated, and red dots represent genes that are differentially upregulated. C GSEA enrichment analysis of transcriptome with high enrichment scores. D Differential binding TFs prediction using the transcriptomic data. E The footprint of top 5 TFs according to the |logFC| was all significantly decreased from young to aged mice genome-wide
Integrated analysis of chromatin accessibility and gene expression during ARHL progressionTo address the potential discrepancy between chromatin accessibility and gene expression levels, a Venn chart analysis was employed to ensure that changes in gene expression were specifically associated with alterations in chromatin accessibility at different gene regions. By focusing on the overlapping regions, we are able to more confidently attribute gene expression changes to specific alterations in the chromatin landscape, providing a more reliable analysis of the regulatory mechanisms underlying age-related changes in gene expression. Nineteen all-up genes and 85 all-down genes were considered directly linked to variations in chromatin accessibility (Fig. 5A, Table S14). In contrast, 20 ATAC-down with mRNA-up genes and 31 ATAC-up with mRNA-down genes would be considered influenced by other factors, such as RNA editing, RNA splicing, and posttranscriptional modifications, rather than changes in chromatin openness. The GO enrichment analysis showed that the assembly and movement of the cilium were necessary for hearing maintenance, reflecting the important role of HCs in ARHL progression (Fig. 5B). The KEGG pathway analysis emphasized the potential role of NF-κB pathway signaling (Fig. 5C).
Fig. 5
Integrated analysis of chromatin accessibility and gene expression during ARHL development. A Venn diagram showing the number of genes in different ATAC and mRNA state, providing a more reliable analysis of the regulatory mechanisms underlying age-related changes in gene expression. B GO enrichment analysis of differentially expressed genes with different peaks. C KEGG analysis of differentially expressed genes with different peaks. D Gene element-based correlation analyses between peak intensity and gene expression, suggesting the importance of chromatin accessibility in the promoter and 5’UTR regions for proximal regulation. R is Pearson’s correlation coefficient and P is the significance test for the correlation coefficient. E The number of genes containing different numbers of peaks in the promoter regions in two groups. Most genes in both groups have 1–3 peaks in their promoter regions. F Comparing expression distribution of up or downregulated genes based on whether they contain promoter peaks or not
To explore the regulatory relationship between chromatin accessibility and gene transcription, we initially conducted an analysis to examine the correlation between the intensity of chromatin accessibility peaks and the expression levels of nearby genes in two distinct groups of mice.
Correlation analysis (Pearson’s correlation coefficient) revealed that peaks in different positions of genes exhibited distinct effects on the regulation of nearby genes. In the control group, the peaks in the intron, exons, and intergenic regions showed the weakest effect on gene expression (intron: R = 0.051; exon: R = 0.04; intergenic regions: R = 0.035; downstream: R = 0.26). The peaks in the 5’UTRs and promoter regions showed a moderate effect on gene expression (5’UTRs: R = 0.36; promoters: R = 0.19, p value < 0.0001) (Fig. 5D). Similar to the case in the aging group, the peaks in the intron, exons, and intergenic regions showed the weakest effect on gene expression (intron: R = 0.032; exon: R = 0.012; intergenic regions: R = 0.026; downstream: R = 0.048), while the peaks in the 5’UTRs and promoter regions showed a moderate effect on gene expression (5’UTRs: R = 0.3; promoters: R = 0.14, p value < 0.0001) (Fig. 5D). The correlation analyses between chromatin accessibility and gene expression suggested the importance of chromatin accessibility in the promoter and 5’UTR regions for proximal regulation, providing insights into the differential regulatory mechanisms associated with distinct chromatin accessibility patterns. We further comprehensively explored the changes in open chromatin in the promoter regions of aging and control cochleae. In control cochleae, 10,124 genes had no peak (n peak = 0), 7456 genes contained a single peak (n peak = 1), and 30,504 genes contained multiple peaks (n peak > 1) in their promoter regions (Fig. 5E). Most genes in both groups have 1–3 peaks in their promoter regions. In aging cochleae, 10,556 genes had no peak (n peak = 0), and 7940 genes contained a single peak (n peak = 1), but 29,715 genes contained multiple peaks (n peak > 1) in their promoter regions. Most upregulated genes containing 0 peaks showed lower expression, while most genes containing peak(s) showed higher expression; most downregulated genes did not contain peaks and showed more obvious fold changes in expression than those with peak(s) (Fig. 5F).
Identification of SASEs and prediction of their target genes in mice with ARHLHistone acetylation plays a crucial role in modulating chromatin accessibility. Acetylated histones can create a more open chromatin structure, facilitating the recruitment of transcription factors and other regulatory proteins to specific genomic regions. To further explore the potential reasons for chromatin accessibility changes in the aging cochleae, we screened the global status of the most common histone acetylation modifications, including H3K9ac, H3K18ac, and H3K27ac. Western blot analysis showed that the levels of H3K27 and H3K18 acetylated histones were decreased in aging cochleae compared to control cochleae (n = 6, Student’s t test, both p < 0.0001), while no significant alteration was detected in H3K9ac (Fig. 6A, B). H3K27ac is a specific histone modification that plays a crucial role in gene regulation and is often found in regions of the genome where gene transcription is highly active. SEs, on the other hand, are large clusters of transcriptional enhancers that drive expression of genes defining cell identity and often physically interact with the promoter region of their target genes, forming chromatin loops and enhancing gene transcription. They are characterized by high levels of H3K27ac and other chromatin marks associated with active transcription, such as mediator complex binding and the presence of transcription factor binding sites [35]. The presence of H3K27ac at SEs reflects their active and highly accessible chromatin state, allowing robust and sustained gene expression. CTCF can bind to specific DNA motifs and create topology associating domains (TADs), which are chromatin loops that restrict the contacts that a promoter makes with distal elements such as enhancers within the loop [36]. The motif analysis indicated that CTCF is the TF most frequently bound to its motif in these samples (i.e., the top 1), also indicating that SEs may play a crucial role in ARHL. Finally, we found that SASEs (senescence-associated SEs) tended to be involved in the process of ARHL development.
Fig. 6
Identification of SASEs and prediction of their target genes in mice with ARHL. A, B Western blot analyses of H3K9ac, H3K18ac, and H3K27ac in the cochleae of 6 W and12 M mice. The statistical significance was represented as * p < 0.05; **p < 0.01; ***p < 0.001;****p < 0.0001. Bar graph results are means ± SD from 6 independent experiments. C The distribution of SASEs region peaks around the transcription start sites and average ATAC-seq signal density in the young state and aging state of cochlea surrounding the SASE regions. D Geneplots of identified SASEs. The position of different peaks was labeled with green cubes and peak numbers. E Geneplots of SASE-linked genes. The position of different peaks and gene exons was marked with green cubes. The thick green line represents the location of SASE
A H3K27ac chromatin immunoprecipitation and sequencing study conducted by Sturmlechner and his colleagues identified 40 common SASEs and 50 transcriptionally activated genes that are associated with these common SASEs in senescent MEFs with senescence induced through three distinct stressors [16]. Other studies have shown that SEs are generally enriched in regions of accessible chromatin; moreover, ATAC-seq has been widely conducted to monitor chromatin accessibility in SE regions and has confirmed that SEs are indeed associated with the phenotype studied in the active or inactive state [37]. First, we drew the peak center plots for all SE region peaks, referring to the known SASE list. As expected, the SE region peaks showed a consistent distribution among the biological replicates, which revealed the high conservation of SASEs related to ARHL (Fig. 6C). The merged plot revealed that the sequencing depth of different SE regions was overall higher in the control group than in the aging group, which indicated that the SASE regions of control cochleae were in a more active and highly accessible chromatin state, allowing the TFs and regulatory factors to bind. In total, we identified 18 SASEs with different peak numbers (n ≥ 1) from the ATAC data and 18 candidate SASE genes. The list of the identified SASE IDs and candidate SASE genes is provided in Table 1. Figure 6D and Figure S5 show the plots of genes associated with candidate SASEs and display the gene plots of SASEs (different peak numbers ≥ 2), which could provide valuable insights into the molecular mechanisms underlying the development and progression of ARHL in mice. The potential functional SE regions of candidate SASE genes were observed in their gene plots, labeled as green squares with peak numbers (Fig. 6E). The identification of SASEs and the prediction of their target genes in ARHL mice could contribute to our understanding of the regulatory networks involved in age-related hearing loss.
Table 1 Identified SASEs related to ARHL with different peaks number and candidate SASE genesChanges in the expression levels of SASE genes in D-gal-induced aging and JQ-1-treated HEI-OC1 cellsThe most common type of ARHL, sensory presbycusis, involves damage to the HCs located at the base of the cochlea. Treating cells or animals with D-gal is a common method used to induce cellular and animal aging for studies of the effects of aging on various biological processes [38]. To investigate the role of candidate SASE genes in the aging of mouse cochlear HCs, we established an aged mouse cochlear HC model by treating HEI-OC1 cells (derived from conditionally immortalized mouse auditory cells) with different D-gal concentrations (2, 5, 10, 15, 20, 30, 40, and 50 mg/mL) for 48 h. CCK8 assay results showed that the cell density began to decrease when the concentration of D-gal was higher than 2 mg/ml, and the cell density was significantly decreased when the D-gal concentration was 10 mg/ml or higher (Fig. 7A, p < 0.01, n = 6). The IC50 of D-gal was 31.77 mg/ml, and 30 mg/ml was considered a suitable concentration to induce cellular aging. DCFH-DA was used to detect the ROS levels in cells with D-gal-induced aging, and fluorescence intensity results showed that the ROS level was significantly increased compared to controls when the D-gal concentration was 30 mg/mL (Fig. 7B). Other hallmarks of aging are decreases in H3, H4, and LaminB1 [39], a series of protein known to contribute to cellular senescence, as indicated by increased C–C motif chemokine ligand 2 (Ccl2) [40], C–C motif chemokine ligand 20(Ccl20) [41], and plasminogen activator inhibitor-1(PAI-1) [42] expression (Fig. 7C). The expression levels of p16 and p21 (both common senescence indicators) increased with increasing D-gal concentration in aging HEI-OC1 cells (Fig. 7D), which indicated that the degree of aging had a positive relationship with increasing D-gal concentration. The four-quadrant analysis for ATAC-seq and RNA-seq showed 185 ATAC-down and mRNA-up genes, 535 ATAC-up and mRNA-down genes, 127 all-up genes and 921 all-down genes (Fig. 7E). We examined the expression-level changes in the top genes in different quadrants between normal and model cells (Fig. 7F) and labeled them in Fig. 7E; the findings were in general accord with our expectations.
Fig. 7
The expression level changes of SASE genes in D-gal-induced aging and JQ-1-treated HEI-OC1 cells. A The CCK8 results showed that the inhibitory rate of HEI-OC1 cells rises as the increased D-gal concentration (n = 6). B The ROS level increased after the HEI-OC1 treated by D-gal (n = 4). C The mRNA level of aging hallmarks in the control and D-gal treated cells (n = 3). D The protein level of p16, p21 increased as the D-gal concentration increased. E The differential expressed genes with different chromatin accessibility state. F The mRNA level changes of the top genes in different four quadrants between normal and mimic-aging cells (n ≥ 4). F The levels of H3K27ac decreased with increasing D-gal concentration. H The mRNA level changes of the SASE-linked genes in D-gal-induced aging cells (n ≥ 3). I The mRNA level changes of the SASE-linked genes after treated by JQ-1(1 μm) (n ≥ 3). J Schematic diagram of possible mechanisms underlying positive role of identified SASEs in ARHL (drawn by Figdraw). During the aging process of the cochlea, the positive regulation of candidate SASE gene expression is inhibited due to decreased chromatin accessibility, and inhibition of SE activity caused by JQ-1 could lead to similar effects. The statistical significance was represented as * p < 0.05; **p < 0.01; ***p < 0.001;****p < 0.0001
Interestingly, we also found that the levels of H3K27ac decreased with increasing D-gal concentration, which revealed that there was overall less chromatin accessibility and less active enhancers in aging cells (Fig. 7G). To investigate the role of SASEs in the aging process of these cells, we evaluated the expression levels of 11 candidate SASE genes. We found that all of them were changed, and 8 of them significantly decreased (Fig. 7H). Based on the above, we hypothesized that the positive regulation of candidate SASE gene expression was inhibited due to decreased chromatin accessibility during the aging process of the cochlea and that the suppression of SE activity could elicit similar responses. SEs are characterized by their larger size, higher transcription factor occupancy, and increased binding of coactivators such as BRD4 and the mediator complex. BRD4, a member of the bromodomain and extraterminal domain (BET) family of proteins, plays a significant role in transcription regulation and chromatin remodeling. JQ-1 is a specific and potent BRD4 inhibitor that can interfere with the recruitment of transcriptional coactivators and other factors necessary for gene expression controlled by active enhancers and SEs [43]. To confirm the regulatory relationship of SASEs with their related genes, HEI-OC1 cells were treated with 1 μM JQ-1 for 48 h. As shown in Fig. 7I, blocking BRD4 led to a slightly greater reduction in the expression level of SE-linked genes compared to that in control cells, including Atp6v1a, Scdcbp, Dcun1d3, Mitf, among others. All eight candidate SASEs and their nearby peaks with different chromatin accessibility are shown in Table 2. Taken together, these data from an aged mouse cochlear HC model reinforce the important roles of candidate SASEs and their related genes in the aging process of HEI-OC1 cells.
Table 2 The chromatin accessibility changes of candidate genes nearby identified SASEs involved in the aging of HEI-OC1 cells (the bold text represents the occurrence within the identified SASE segment)
Comments (0)