
Remember me









Hepatic encephalopathy (HE) is a common decompensation in cirrhosis that is associated with increased mortality and frequent readmissions (1,2). Cirrhosis and HE are associated with high ammonia levels, which may be affected by the microbiome, diet, and glutaminase enzyme activity (3–6). Moreover, changes in ammonia levels after defined protein loads have prognostic value, and there has long been interest in the role of diet in managing HE symptoms (7–9).
Most Western countries follow a diet that is rich in simple carbohydrates and red meat and low fiber, which could enhance ammoniagenesis. Some studies have evaluated the utility of a vegetarian diet in patients with HE with a potential for beneficial effects on ammonia levels and mental status (10). However, changes in the entire diet are difficult to achieve and sustain even if proven to be beneficial and require extensive behavioral changes (11). Therefore, the diet occasionally to add alternatives to meat products could be an option. The role of protein source substitution needs to be investigated, but the impact of meal substitution against the background of a Western non-vegetarian diet in cirrhosis is unclear. We aimed in a randomized clinical trial to determine the impact of substituting one meal with equal protein-containing vegan, vegetarian, or meat protein on ammonia levels, microbiome, and metabolomics in outpatients with cirrhosis who typically follow a Western non-vegetarian diet.
METHODS Study populationOutpatients with cirrhosis who were older than 18 years consuming a stable Western, non-vegetarian diet were recruited after obtaining informed consent. If patients had prior HE, only those on stable treatment for more than 6 weeks were included. In addition, patients needed to be able and willing to give consent and did not have known allergies to ingredients in planned meals. We excluded patients with body mass index greater than 40 or less than 18.5, Model for End-stage Liver Disease sodium (MELD-Na) greater than 23, prior transjugular intrahepatic portosystemic shunt, unclear diagnosis of cirrhosis, food allergies, recent antibiotic or probiotic use in the past 6 weeks, not on stable HE treatment, not on a stable Western non-vegetarian diet, or those with current valproate or corticosteroid use. The trial was registered and approved by the institutional review board.
Study designBaseline dietary history, cirrhosis details, MELD-Na, and baseline characteristics were collected from all participants after obtaining informed consent (Figure 1a). Patients were instructed to fast overnight and were randomized 1:1:1 into meat, vegan, or vegetarian meals using a random number generator. After overnight fasting, patients ate the entire meal under observation and had blood drawn at baseline and hourly for 3 hours after the meal. Blood obtained for serum ammonia analysis were collected on ice, which was immediately brought to the VA laboratories for analysis with a reference range of 11–32 μmol/L. No other food or drink was allowed after meal, and all subjects were observed for HE.
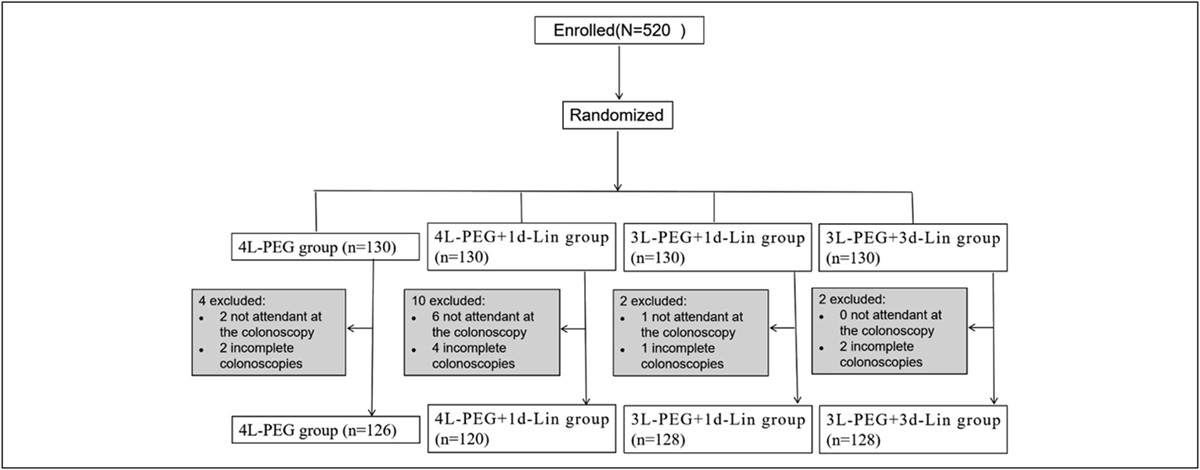
 Figure 1.:
Figure 1.: Study flow and CONSORT diagram. (a) Flowchart of the study design: subjects were randomized in a 1:1:1 fashion to meat, vegan, and vegetarian meals, and blood was drawn hourly for 3 hours after meal for serum ammonia and metabolomics. Stool was collected at baseline for microbiome analysis. (b) CONSORT diagram: There were no dropouts after consent, and all subjects were included in the analysis. CONSORT, Consolidated Standards of Reporting Trials.
Meal contentsPatients were given meat, vegetarian, or vegan burger meal with equal amounts of protein (20 g). We chose 20 g since it is reflective of traditional protein consumption in Western meals and has been tested in our population before (12). All meals contained low-fat potato chips, a whole-grain bun, and water without any condiments (see Supplementary Digital Content, Table S1, https://links.lww.com/CTG/B119). To ensure protein quantities were equal, the patties were weighed and portions adjusted by a registered dietician (VT).
Statistical analysis Sample size.A prior study using 20 g of non-meat protein resulted in 12% increase in ammonia over baseline at 1 hour with SD of 4% (12). Assuming that the change in ammonia with meat would be double (i.e., 24%) and a power of 80% and that vegan and vegetarian meals would behave similarly, we would need 4 subjects per arm. However, since we also included patients with prior HE, we allowed for 10 subjects per arm (5 with and 5 without prior HE).
Sample analysis and statistics.Baseline characteristics between the 3 groups were evaluated using parametric (analysis of variance [ANOVA]) and non-parametric (Kruskal-Wallis) tests as appropriate. Overall, a 2-way repeated-measures ANOVA (RMANOVA) for time, diet, and interaction was performed for serum ammonia levels. Then, we performed one-way RMANOVA within groups for ammonia and ANOVA between each time point for ammonia. Stool and serum microbiome were analyzed using published techniques (13,14). Stool collected at baseline and serum collected at all 4 time points were analyzed. Briefly, stool was stored in RNA later and then extracted using published techniques. Stool 16S ribosomal ribonucleic acid (16SrRNA) analysis was performed, with analysis of α and β diversity at baseline between groups. Differences in bacterial relative abundances at the genus/family level at baseline between groups were analyzed using linear discriminant analysis effect size (15). Circulating nucleic acids were extracted from blood plasma using a column-based QIAamp Circulating Nucleic Acid Isolation Kit (55,114; Qiagen) according to the manufacturer's protocol (14). Specific primer pair sets 27 F and 338 R targeting the hypervariable region V1-V2 were used in the polymerase chain reaction process. Sequencing of the 16S rRNA gene was performed using the Illumina MiSeq platform (Illumina) and a dual-indexing approach. The acquired sequencing data were assigned into amplicon sequencing variants and taxonomically annotated against the ribosomal database project V16 (RDP v16) using the “dada2” software package in R. Details are provided in the Supplementary Digital Content (see Supplement, https://links.lww.com/CTG/B119).
Metabolomics were performed as per prior published studies (16). Briefly, Metabolon's liquid chromatography-mass spectrometry platform was used to analyze serum, and changes over baseline and at hourly intervals within and between groups were analyzed. Following imputation of missing values, if any, with the minimum observed value for each compound and log transformation, 2-way/repeated-measures ANOVA was used to identify biochemicals that differed significantly between experimental groups. Analysis by 2-way ANOVA with repeated measures identified biochemicals exhibiting significant interaction and main effects for experimental parameters of treatment and time point. Two-way RMANOVA was performed for time and meals for metabolomics. Details are provided in the Supplementary Digital Content (see Supplement, https://links.lww.com/CTG/B119).
RESULTS DemographicsForty-four patients were considered, 6 were not interested, 4 were not consistently on a Western non-vegetarian diet, and 3 did not have a clear diagnosis of cirrhosis, and one person had a food allergy. Ultimately, 30 men were included in this study, 50% of whom had prior HE and were evenly distributed across treatment arms (Table 1, Figure 1b). The patients were recruited between June 2022 and December 2022. There were no statistically significant differences in age, body mass index, MELD-Na, etiology of cirrhosis, prior ascites, or variceal bleeding among groups. There were no significant differences in lactulose, rifaximin, proton pump inhibitor, or statin use among the 3 groups.
Table 1. - Clinical and biochemical characteristics of the study population Subject characteristics Meat (n = 10) Vegan (n = 10) Vegetarian (n = 10) Age 66.2 ± 2.7 70.5 ± 3.1 66.1 ± 7.8 Male sex 10 10 10 BMI 32.2 ± 4.3 29.2 ± 5.8 29.9 ± 6.9 Diabetes 5 (50%) 6 (60%) 5 (50%) On insulin 2 (20%) 2 (20%) 1 (10%) Cirrhosis details Etiology (HCV/Alcohol/MASH/others) 4/4/2/0 2/5/3/0 2/5/3/0 Prior HE 5 (50%) 5 (50%) 5 (50%) Prior ascites 5 (50%) 4 (40%) 4 (40%) Prior variceal bleeding 2 (20%) 1(10%) 1(10%) Diuretic use 4 (40%) 4 (40%) 3 (3%) Lactulose 5 (50%) 5 (50%) 5 (50%) Rifaximin 4 (40%) 4 (40%) 3 (30%) Proton pump inhibitor 7 (70%) 7 (70%) 8 (80%) Statin 4 (40%) 5 (50%) 5 (50%) Laboratory values MELD-Na 10.1 ± 4.5 8.9 ± 2.9 9.1 ± 3.0 INR 1.24 ± 0.35 1.06 ± 0.12 1.16 ± 0.21 Serum creatinine 1.03 ± 0.57 1.18 ± 0.37 1.03 ± 0.25 Serum bilirubin (mg/dL) 1.03 ± 0.81 0.75 ± 0.34 0.51 ± 0.27 Serum sodium (Meq/L) 139.7 ± 2.1 139.8 ± 3.8 138.5 ± 3.3 WBC count 5.28 ± 1.73 5.55 ± 2.82 5.30 ± 1.16 Hemoglobin 13.20 ± 0.88 13.43 ± 1.63 13.71 ± 1.28 Platelet count 147.2 ± 105.8 177.6 ± 95.7 147.3 ± 64.8 ALT 30.8 ± 15.4 34.3 ± 12.7 36.5 ± 12.7 AST 36.6 ± 18.7 29.7 ± 14.2 35.6 ± 12.6 Alkaline phosphatase 103.1 ± 38.1 101.7 ± 73.7 105.3 ± 33.8ALT, alanine transaminase; AST, aspartate transaminase, BMI, body mass index; HCV, hepatitis C; INR, international normalized ratio; MASH, metabolic-associated steatohepatitis; MELD-Na–Model for End-stage Liver Disease sodium.
Dietary intake over the preceeding 3 days before intervention showed statistically similar intake of red meat, poultry, fish, eggs, bread, cheese, rice, fruits, vegetables, yogurt, coffee, tea, carbonated caffeinated, and decaffeinated beverages among the 3 groups (see Supplementary Digital Content, Table S2, https://links.lww.com/CTG/B119). All subjects completed the entire meal, and none of the patients developed HE symptoms during the 3-hour observation.
Serum ammoniaSerum ammonia at baseline was similar among the 3 groups. Serum ammonia significantly elevated in the meat group compared with baseline, but not in the vegan or vegetarian group (Table 2, Figure 2a). Between groups, there was little difference at baseline but higher ammonia levels were seen at 1 and 2 hours after meal, whereas on the other hand the only group that showed increase over baseline using one-way RMANOVA was the meat group (Table 2). We also performed a 2-way RMANOVA, where time was significant (P = 0.043, F = 2.389) as was the interaction term between time and diet (P = 0.0009, F = 4.251) with a trend toward change in diet (P = 0.09, F = 2.596, Figure 2a). On post hoc analysis, this was driven by the meat-assigned group. Patients with prior HE had higher serum levels of ammonia regardless of treatment group at all time points (Figure 2b–d). Trend in ammonia was similar in the meat group in both patients with and without prior HE.
Table 2. - Changes in Ammonia over time Ammonia mean (SD) Meat (n = 10) Vegan (n = 10) Vegetarian (n = 10) P value between groups (ANOVA) Baseline 58.2 (46.2) 36.1 (14.9) 52.9 (36.0) 0.20 1 hr post 75.1 (43.8)a 39.3 (14.8) 50.0 (25.4) 0.03 2 hr post 76.4 (49.9)a 41.2 (12.5) 47.8 (22.9) 0.04 3 hr post 74.1 (58.6)a 41.5 (36.0) 45.5 (27.5) 0.12 One-way RMANOVA within groups (P value) 0.001 0.33 0.32 Tests between groups are one-way ANOVA while within group are one-way RMANOVA. 2-way ANOVA analyses are shown in Figure 2a.ANOVA, analysis of variance; RMANOVA, repeated-measures ANOVA.
aP < 0.05 between meat and other groups.
 Figure 2.:
Figure 2.: Serum ammonia changes from baseline in meat, vegan, and vegetarian groups. (a) There was a statistically significant increase in serum ammonia from baseline at each hourly time point in the meat group but not in the vegan or vegetarian group using repeated-measures ammonia analysis. (b–d) Changes in serum ammonia based on prior hepatic encephalopathy or no prior hepatic encephalopathy. There was a similar trend in ammonia with prior HE vs no prior HE with no statistically significant differences in either the meat, vegan, or vegetarian meals. Red: HE, Black: No-HE. HE, hepatic encephalopathy.
Stool microbiomeStool microbiome was similar across groups without changes in α or β-diversity metrics (Figure 3). We also did not find any bacterial change differences at the genus or family level on linear discriminant analysis effect size.
 Figure 3.:
Figure 3.: Baseline stool microbiome. (a–c) Alpha diversity values of stool microbiome at baseline. (d) Beta diversity of stool microbiome at baseline. No significant differences were seen.
Serum microbiomeThere was low yield with contamination in plasma microbiome, with predominant Cutibacterium spp belonging to Actinobacteria (see Supplementary Digital Content, https://links.lww.com/CTG/B119). The water control also had similar features. Therefore, this was not analyzed further.
MetabolomicsSince ammonia changes were higher in meat vs others, we focused the metabolomic analysis on meat vs vegan and meat vs vegetarian. The serum data set comprised a total of 1,499 biochemicals, 1,189 compounds of known identity (named biochemicals), and 310 compounds of unknown structural identity (unnamed biochemicals). Compared with the vegetarian group, 97 metabolites were different in the meat group, of which 30 were higher and 67 were lower. Similarly, compared with the vegan group, 89 metabolites were significantly different, 65 were lower and 24 were higher. In patients with HE, 71 metabolites were different in the vegetarian compared with meat group with 26 higher and 45 lower while in the vegan group, 106 metabolites were different vs meat (55 higher and 51 lower).127 metabolites were different in all subjects based on treatment and time point interactions while 86 were different in the patients with HE alone. Data are presented in Supplementary Digital Content (see Figure S2-5, https://links.lww.com/CTG/B119) and Tables 3 and 4.
Table 3. - Metabolomic changes at 1 hour based on analysis of variance contrasts in all subjects Pathway Metabolite (meat/vegetarian fold change) Metabolite (meat/vegan fold change) Higher in meat vs other Food constituents 5-hydroxylysine—connective tissue (2.06), Histidylalanine—skeletal muscle (4.00) 5-hydroxylysine—connective tissue (2.19), Histidylalanine—skeletal muscle (2.61), Acesulfame—sweetener (13.63) Lipids sphingolipids lactosyl-N-behenoyl-sphingosine (1.51), palmitoyl sphingomyelin (1.22), glycosyl ceramide (2.56), palmitoyl-sphingosine-phosphoethanolamine (1.3) lactosyl-N-behenoyl-sphingosine (1.63), palmitoyl sphingomyelin (1.18), glycosyl ceramide (2.41), palmitoyl-sphingosine-phosphoethanolamine (1.25) Lipids: others cis-3,4-methyleneheptanoate (1.53) cis-3,4-methyleneheptanoate (1.66), cortolone glucuronide (1.83), cholesterol sulfate (1.33), glucuronide (1.83) Nucleoside Beta-alanine (1.55) Beta-alanine (1.68), Cytidine (1.47) Xenobiotic (2,4 or 2,5)-dimethylphenol sulfate (1.86), methylnaphthyl sulfate (2.44), 2-ethylphenylsulfate (3.45) (2,4 or 2,5)-dimethylphenol sulfate (6.06), methylnaphthyl sulfate (3.05), 2-ethylphenylsulfate (9.86), methyl-4-hydroxybenzoate sulfate (4.72) Lower in meat vs other Food constituents Genistein sulfate (0.31) (soy), Daidzein sulfate (0.14) (soy), Indolin-2-one (0.40) Genistein sulfate (0.52) (soy), Daidzein sulfate (0.32) (soy), Maltol sulfate (0.20) (flavoring agent), Allin (0.73) (garlic), S-allylcystein (0.23) (garlic), N-acetyl-S-allyl-cysteine (0.17) (garlic), Indolin-2-one (0.49), 2-acetamidophenol sulfate (0.25), O-acetyl-homoserine (0.38) Branched chain amino acid metabolites 4-methyl-2-oxopentanoate (0.55), 3-methyl-2-oxovalerate (0.57), Isovaleryl-carnitine (0.55), isobutyryl-carnitine (0.42), isobutyryl-glycine (0.58) 4-methyl-2-oxopentanoate (0.58), 3-methyl-2-oxovalerate (0.63), 3-methylglutaconate (0.5), tigloyl-glycine (0.46), ethyl-malonate (0.62), isovaleryl-carnitine (0.58), isobutyryl-carnitine (0.32), isobutyryl-glycine (0.4) Tyrosine metabolites Dopamine 4-sulfate (0.63), dopamine 3-O-sulfate (0.65) Urea cycle Ornithine (0.77) Glycine, metabolism N-acetyl-glycine (0.49) Methionine, cysteine, SAM and taurine metabolism Cysteine (0.79) Cysteine (0.76), S-methylcysteine sulfoxide (0.61) Lysine metabolism N2-acetyl,N6-methyllysine (0.31), N2-acetyl,N6,N6-dimethyllysine (0.18), N6,N6-dimethyllysine (0.58) Lipid Propionyl glycine (0.25) Lysophopsholipids: 1-linolenoyl-GPC (0.7), 1-dihomo-linolenoyl-GPC (0.64), 1-dihomo-linolenoyl-GPE (0.51),1-arachidonoyl-GPE (0.62),1-palmitoleoyl-GPC (0.60), 1-palmitoleoyl-GPC (0.7), 2-oleoyl-GPC (0.69), 1-adrenoyl-GPC (0.69), Others: 1-oleoyl-2-arachidonoyl-GPE (0.51), sebacate (0.47), 2-myristoylglycerol (0.56), Butyrylcarnitine (0.60) Vitamin A Retinal (0.64) Retinal (0.58) Xenobiotic 4-hydroxychlorothalonil (0.38), 4-acetamidobenzoate (0.46), perfluorooctanoate (0.53), perfluorohexanesulfonate (0.5)GPC, glycerophosphocholine; GPE, glycerophosphatidylethanolamine; SAM, S-Adenosyl methionine.
GPA, glyceryl phopsphate; GPC, glycerophosphocholine; GPE, glycerophosphatidylethanolamine; SAM, S-Adenosyl methionine; TCA cycle, the citric acid cycle.
On repeated-measures analysis with both time and meal interactions, markers of food consumption histidyl-alanine and 5-hydroxylysine were higher, which are associated with skeletal muscle and connective tissue consumption, respectively, while daidzein sulfate and genistein sulfate, both markers of soy consumption were lower in the meat compared with vegetarian or vegan-randomized groups. Compared with vegan, meat patients were lower in O-acetylhomoserine and S-allylcysteine. Compared with meat vs vegetarian, only eugenol sulfate was lower in meat-assigned groups. Tyrosine metabolites (Dopamine 3 and 4-sulfate) were lower in meat vs vegan-assigned patients (Figure 4).
 Figure 4.:
Figure 4.: Summary of notable differences in the metabolomic profile comparing meat vs vegetarian/vegan meals. Metabolites higher in the meat group are colored red while those that were lower in the meat group compared with the vegetarian/vegan group are colored green. GPC, glycerophosphocholine; GPE, glycerophosphatidylethanolamine; TCA cycle, the citric acid cycle.
Differences with time only on metabolomics.When only time was considered on the repeated-measures analysis for patients with HE, the meat group had higher lipid metabolites compared with both vegan and vegetarian. Meat-assigned patients also showed higher amino acids associated with HE and ammoniagenesis (methionine and 5-hydroxylysine) than the vegan group. The meat group had higher phospholipid/ceramide, methionine, and 5-hydroxylysine levels than the vegan group. The meat group had higher long-chain fatty acid, sphingolipid, acylcarnitine metabolites, and phosphatidylcholine than the vegetarian group, but lower lysophosphatidylcholine than the vegan or vegetarian group. The vegan group had higher N-acyl amino acid levels than the meat or vegetarian group.
Patients with prior HE on metabolomics.In patients with prior HE only, on repeated measures for time and meal interactions, 5-hydroxylysine was higher and 3-hydroxy-2-methylpyridine sulfate was lower in meat vs other groups. Comparing meat with vegan, O-acetylhomoserine, S-allylcysteine, dopamine 4-sulfate, 4-methylguaiacol sulfate, and 4-methylcatechol sulfate were lower in meat. No major changes between the meat and vegetarian groups were seen.
When comparisons at time points were considered, methionine metabolomic moieties were higher in meat (cysteine, sulfinic acid, methionine) while branched chain amino acids (BCAA) (3-methylglutaconate, isobutrylcarnitine, isobutrylglycine, methylsuccinate, N-acetylleucine, methylsuccinoylcarnitine), tyrosine (gentisate, phenol sulfate), and tryptophan (3-indoxyl sulfate) were lower in the meat group compared with the vegan group.
DISCUSSIONIn this randomized clinical trial, we found that substitution of just one meat-based meal with a vegan or vegetarian alternative was associated with decreased postprandial ammonia and may improve levels of serum metabolites that are associated with ammoniagenesis and HE. Serum ammonia was elevated compared with baseline in the meat group at all time measurements but not in the vegan or vegetarian groups. Our results also show differences in metabolites that may have implications for the development of HE or ammoniagenesis, and given that subjects had similar microbiome at baseline, this is likely best explained by diet. Notable differences between the groups were differences in amino acid metabolites with less branched chain amino acid metabolites in the meat group, which is less ammoniagenic. There was also an increase in metabolites associated with the urea cycle; notably acylcarnitines were higher in the vegetarian and vegan groups than the meat group. Lipid profiles were also different in the meat group having higher sphingolipids, and lower lysophospholipids with altered lipid metabolism has been identified in patients with advanced liver disease.
The results in our study in regard to reduced ammonia when comparing a non-meat-based protein diet with a meat-based diet (10,17,18). To our knowledge, this is the first study to analyze the effects of ammoniagenesis after a full meal rather than an isolated protein load. Prior studies analyzed the effect of vegetarian diet over prolonged periods, noting improved serum ammonia after several days of dietary intervention (10,19) Our results show that this decrease in ammoniagenesis can occur with substitution of just one meal. These results are promising as dietary interventions can be difficult to sustain in the long term and patients accustomed to a Western diet may find transitioning to a full vegan or vegetarian diet challenging (11).
Through metabolomic analysis, we found potential mechanisms for the decreased serum ammonia concentration in the vegetarian and vegan groups that needs to be carefully interpreted as a proof of concept given the overall small sample size of the trial. There was a decrease in branched-chain amino acid metabolites such as 4-methyl-2-oxopentanoate and 3-methyl-2-oxovalerate as well as a decrease in acylcarnitines, which are involved in BCAA metabolism in subjects who had the meat-containing meal compared with those who received either the vegetarian or vegan meals. Not only are BCAAs less ammoniagenic but the metabolism of branch chain amino acids can have a positive effect on ammonia clearance through the formation of glutamate, which in turn can incorporate a free ammonia molecule to produce glutamine (20,21). BCAA supplementation has often been proposed as a potential treatment option for patients with HE (22).
While the specific amino acid profile of the meals provided were not analyzed, prior studies have compared amino acid make-up of different protein sources in foods. Notably, methionine and lysine tend to be lower in plant-based protein sources than animal sources such as whey and egg (23). When the amino acid content of different patties of beef, soy, or pea protein, the plant-based proteins tended to be lower in cysteine and methionine (24). Differences in glutamine, which has an important role in ammoniagenesis by the gut, was not able to be determined in the protein sources due to its conversion to glutamic acid in processing (25).
Also notable was the decrease in acylcarnitines in the meat group compared with the vegetarian and vegan groups. Not only are acylcarnitines involved in the metabolism of BCAAs but are also important in ammonia excretion via the urea cycle. Acylcarnitines facilitate passage of lipids into the mitochondria for beta-oxidation, producing acetyl-CoA, a necessary substrate for the formation of n-acetylglutamate, activating the rate limiting step of the urea cycle (26). Non-cirrhotic hyperammonia-associated conditions such as urea cycle defects or valproate toxicity are associated with acylcarnitine depletion, and carnitine supplementation is often used as a treatment in these conditions (27,28).
There was noted to be an altered lipid profile in patients who had the meat-containing meal compared with those who did not. Notably, sphingomyelins elevated in the meat group but not in the vegetarian or meat group while lysophospholipids were decreased. Alterations in the lipidomics of patients with liver disease have been previously noted with an increase in sphingomyelins and ceramides and a decrease in phospholipids such as phosphatidylcholine and phosphatidylethanolamines associated with more advanced liver disease (29–32). A prior study analyzing the lipid profile of patients with minimal HE found an increase in sphingomyelins and a decrease in lysophospholipids (33). An increase in ceramide may provide an additional link to ammonia production through its ability to decrease fatty acid metabolism through inhibition of the electron transport chain (34).
The sources and flux of circulating ammonia in health and in cirrhosis are complex (5,35). Diet, gut microbiota, expression of small bowel transglutaminase, and liver/muscle health are involved (5,35). In our study, we found a similar composition of stool microbiome at baseline and otherwise similar clinical and HE-related characteristics in our patient population. The circulating microbiome analysis did not provide meaningful answers due technical reasons, which are often seen in low-biomass fluids (36). While we did not assess sarcopenia formally, we do not anticipate ammonia changes over 3 hours to be reflective of muscle function in this stable outpatient population (37). Given the accompanying changes in metabolomic profiles and differences in meal composition, the most likely source of ammonia change here is diet related.
Dietary changes can be difficult to maintain, but there are increasing data that changes in the microbiome can facilitate dietary preferences. In a recent study, overweight subjects were given a prebiotic of inulin, which resulted in more short-chain fatty acid (SCFA) producing bacteria with accompanying changes in the brain activation related to food preferences (38). The implications of this study go beyond just those of food decision making with HE, as SCFA such as butyrate is shown to maintain gut integrity and is protective against HE (39). Other studies have shown an association of high meat consumption with a decrease in SCFA producing gut bacteria such as Lachnospiraceae and Ruminococcaceae (40).
Findings of our study are limited due to a relatively small sample size, studying the impact of only one meal, lack of clinical outcomes, and no formal sarcopenia assessment. Potential mechanistic analysis using metabolomics also need to be interpreted as a proof of concept since multiple corrections adjustment in this small cohort was not applied. However, given the prognostication of ammonia increases after protein loads, our findings would be potentially informative of a larger trial (41,42). Another potential limitation of our study is that the 3-hour time frame of our study may not be sufficient to capture all meaningful differences in the serum metabolites, especially in patients with impaired synthetic function. We also did not perform cognitive testing or urine collection. A larger trial is needed to confirm if meal substitution with vegan/vegetarian alternatives protects against development of HE. Caution should be implemented in recommending a non-meat-based diet to patients with HE that it does not come at the expense of decreased oral protein intake (43). Cirrhosis is a catabolic state, and a low protein diet is associated with sarcopenia, which is detrimental to nitrogen balance as well as associated with mortality (44).
In summary, substitution of one meat-based meal with a vegetarian or vegan alternative resulted in decreased ammonia production regardless of prior HE, and differences in the metabolic profile were noted. Intermittent substitution of a vegetarian or vegan meal in place of a meat-based meal could be an option in patients who usually follow a meat-based diet. Larger studies with longer duration should focus on whether intermittent vegetarian or vegan meals would result in clinically meaningful outcomes in at-risk patients, such as prevention of HE in patients with cirrhosis who normally follow a Western meat-based diet and should focus on appropriate patient selection.
CONFLICTS OF INTERESTGuarantor of the article: Jasmohan S. Bajaj, MD, MS.
Specific author contributions: J.S.B. was involved in all aspects of the study. B.D.B. was involved in research conduct and drafting the manuscript. V.T. was the nutritionist who su
Comments (0)