
Remember me
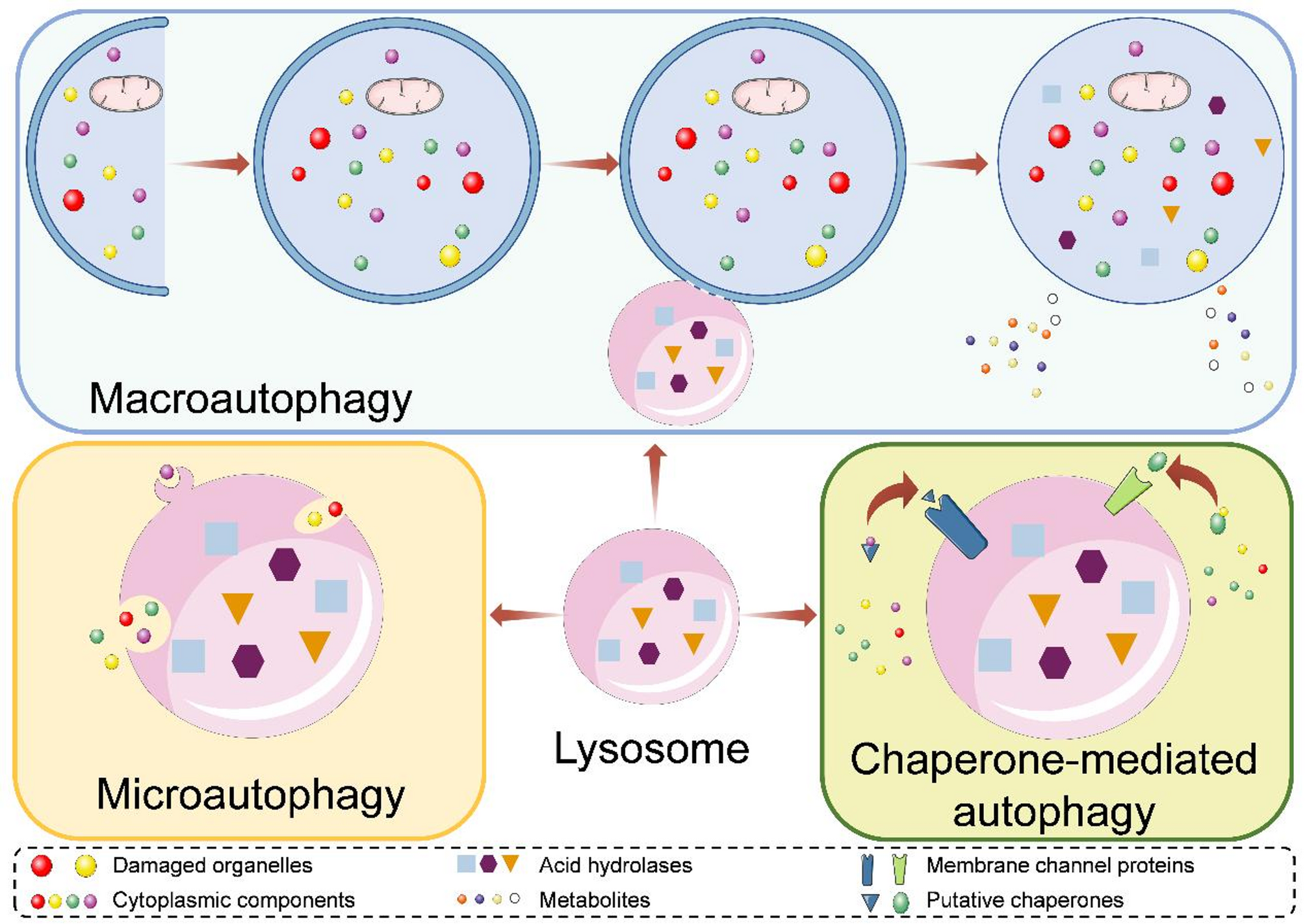




Macroautophagy (henceforth referred to as ‘autophagy’) serves as an important adaptive mechanism to mechanical forces in PDL cells and tissues. During OTM, autophagy is activated and Atg proteins are increased in vitro and in vivo. Lü et al. have established a rat OTM model and have found that Beclin-1 (BECN1) and microtubule-associated protein 1 light chain 3 (MAP1LC3, LC3, also known as Atg8 in yeast) expressions are elevated, with osteoclasts beginning to increase after 1 d on the compression side (Lü et al. 2019). Then, they further noticed that autophagy markers and tumor necrosis factor-alpha (TNF-α) expressions fluctuate but increase with a similar trend under orthodontic force (Xu et al. 2020). Further, autophagy is rapidly initiated and apoptosis is gradually increased after force loading (Wang et al. 2021). Thus, the initiation of autophagy may be associated with osteoclastogenesis, apoptosis, and TNF-α upregulation during OTM. Besides, regulating autophagy affects OTM. Rapa rescues OTM-induced bone density decline and inflammation increase to alleviate OTM, while 3-MA has reverse results in murine OTM experiments with intraperitoneal injection of 3-MA and Rapa (Chen and Hua 2021). Accumulating evidence has revealed that autophagy regulators contribute to OTM and OIIRR (Table 1).
CF and TF are orthodontic forces to determine the speed of OTM, and FSS is another major force in OTM. We summarize the roles of autophagy in OTM based on different forces in Table 2. In addition, hypoxia usually occurs after orthodontic force application. Therefore, we focus on the activities of autophagy in hypoxia, CF, TF, and FSS in OTM-related cells (Fig. 4) and the underlying mechanisms of OTM-related processes (Fig. 5).
Table 2 Roles of autophagy in OTM-related forcesFig. 4
Roles of autophagy in OTM. Under CF, autophagy is activated in most OTM-related cells except for cementoblasts, with inconsistent results of osteoclast autophagy. TF, FSS, and hypoxia activate autophagy in studied cells
Fig. 5
Effects and mechanisms of autophagy in OTM. CF inhibits autophagy to suppress cementogenesis but activates autophagy to promote osteogenesis, apoptosis, and M1 polarization and to impede cell migration. CF differently regulates autophagy and affects osteoclastogenesis. TF induces autophagy to enhance osteogenesis, cementogenesis, spherical change, and anaerobic oxidation, but to decrease cell death and osteoclastogenesis. Hypoxia enhances autophagy to inhibit osteogenesis but increases osteoclastogenesis and apoptosis CF, compressive force; TF, tensile force; CMS, Cyclic mechanical stretch; CTS, cyclic tensile strain/cyclic tensile stress; STS, static tensile strain; SMS, static mechanical stretch; FoxO3, Forkhead box O3; IL-6, Interleukin 6; MTORC2, mechanistic target of rapamycin complex 2; TFE3, transcription factor E3; NLRP3, nucleotide-binding domain (NBD), leucine-rich repeat (LRR), and pyrin domain (PYD)-containing protein 3; BECN1, Beclin-1; p-AKT, phosphorylation of protein kinase B; MMP9/13, matrix metalloproteinase 9/13; YAP, Yes-associated protein; ULK1, unc-51 like autophagy activating kinase 1; AMPK, adenosine monophosphate-activated protein kinase; p-ERK1/2, phosphorylation of extracellular signal-regulated kinase 1/2
Autophagy in OTM-related hypoxiaOrthodontic force occludes the periodontal vessels on the compression side, resulting in ischemia and hypoxia (He et al. 2023). Hypoxia is a known autophagy inducer in human diseases (Fu et al. 2024). A transcriptome profile of cementoblasts indicates that hypoxia-related genes are upregulated under CF (Liu et al. 2019). Moreover, a proteomic analysis uncovers autophagy in human PDLCs under hypoxia (Li et al. 2019). Therefore, studying autophagy of OTM under hypoxic conditions may provide a theoretical basis for elucidating OTM mechanisms (Table 3 and Fig. 5).
Table 3 Roles of autophagy in OTM-related hypoxiaHypoxia-induced autophagy represses osteogenic differentiationCobalt (II) chloride (CoCl2)-induced chemical hypoxia is one of the most commonly used models for simulating hypoxia (Li et al. 2022b). CoCl2 significantly increases the expression of a circular RNA (circRNA) has_circ_0003489 located at the gene for cyclin-dependent kinase 8 (circCDK8) (Zheng et al. 2020). In addition, CoCl2 induces autophagy and apoptosis, and inhibition of autophagy promotes osteogenesis in CoCl2-treated PDL stem cells (PDLSCs). Additionally, overexpression of circCDK8 prompts autophagy and apoptosis through mTOR signaling, while silencing circCDK8 reverses the inhibitory effects of CoCl2 on the osteogenic differentiation of PDLSCs. Interestingly, osteoblast-related markers are increased in CoCl2-treated PDLSCs at 6 and 12 h but decreased at 24, 48, and 72 h, but the authors don’t explore it in depth, which may not be their focus but still a research topic. In conclusion, circCDK8 represses the osteogenesis of PDLSCs by triggering autophagy under hypoxia, which indicates that CircCDK8 is a new target for intervention in OTM.
The optimal hypoxia model involves using a low oxygen concentration in a hypoxia chamber or a Tri-gas incubator. In 2023, Xu et al. utilize a Tri-gas incubator with 5% O2 and find that hypoxia increases autophagy, Fork-head box protein O1 (FOXO1), and Histone deacetylase 6 (HDAC6), but suppresses osteogenesis in rat bone marrow-derived mesenchymal stem cells (BMMSCs), which is verified by the unilateral nasal obstruction (UNO) rat model (Xu et al. 2023). Further analysis reveals that HDAC6 knockdown alleviates hypoxia-diminished osteogenesis, and FOXO1 knockdown attenuates hypoxia-inhibited osteogenesis by elevating autophagy. However, it remains unclear whether FOXO1 directly downregulates osteogenic differentiation, and it is unclear whether HDAC6 or FOXO1 plays a more significant role in osteogenesis. Furthermore, it needs to be verified whether the roles of hypoxia-induced autophagy in osteogenic differentiation are similar in other tissues.
Hypoxia induces osteoblast autophagyExcept for stem cells, a 1% O2 hypoxia environment reduces osteoblast proliferation but activates autophagy (Zhang et al. 2019). To investigate the regulatory pathway, researchers further discovered that hypoxia increases the levels of nicotinamide adenine dinucleotide phosphate (NADPH) oxidase 5 (NOX5) and NADPH oxidase subunit p22phox (He et al. 2021). Subsequently, they observed that inhibition of NOX5 or p22phox by siRNAs decreases autophagy, ROS, and apoptosis under hypoxia conditions, thereby promoting osteoblast proliferation. However, this study only validated the roles of p22phox and NOX5 in vitro, and further in vivo experiments are needed. In addition, the specific molecular relationship between osteoblast hypoxia-induced autophagy and ROS also needs to be further studied. Nevertheless, these two studies demonstrate that hypoxia induces osteoblast autophagy, providing a basis for future studies of OTM-induced changes in hypoxic environments.
Autophagy attenuates hypoxia-induced apoptosis in cementoblastsIn addition to osteoblasts, cementoblast apoptosis is also increased under OTM-decreased periodontal oxygen partial pressure, which may be the mechanism of OIIRR (Mizoguchi et al. 2017). As a nitric oxide donor, L-arginine partly protects against hydrogen peroxide-induced damage by improving mitochondrial function and alleviating cellular apoptosis and autophagy (Zhang et al. 2021a). In 2022, Xu et al. demonstrated that L-arginine inhibits hypoxia-induced apoptosis while increasing Sirtuin 1 (Sirt1) expression in OCCM-30 cells (Xu et al. 2022b). Furthermore, Resveratrol increases autophagy and reduces apoptosis, whereas autophagy inhibition attenuates Sirt1-decreased apoptosis. In vivo, L-arginine upregulates Sirt1 and activates autophagy, thereby reducing OIIRR in rats. Collectively, L-arginine reduces cementoblast apoptosis under hypoxia and attenuates OIIRR in rats, which may be potentially achieved through by Sirt1-enhanced autophagy. The authors interpret Resveratrol as an inducer of autophagy, but they could have applied L-arginine directly if they wanted to clarify its role in autophagy. Notably, the authors acknowledge that clarifying the roles of L-arginine on inflammation is meaningful for understanding OIIRR, which is considered a complex sterile inflammatory response.
Autophagy enhances hypoxia-induced osteoclastogenesisOsteoclasts are another crucial cell type mediating bone resorption. 0.2% oxygen hypoxia enhances autophagy and osteoclastogenesis in RAW 264.7 cells (Zhao et al. 2011). Moreover, hypoxia-induced osteoclastogenesis is attenuated after inhibiting autophagy by ATG5 knockdown or 3-MA application. Furthermore, hypoxia-induced autophagy is caused by upregulating hypoxia-inducible factor-1 alpha (HIF-1α)-dependent Bcl2 adenovirus E1B 19 kDa interacting protein 3 (BNIP3), and knocking down of HIF-1α or BNIP3 alleviates hypoxia-induced autophagy and osteoclastogenesis. They also observe that the number of surviving cells increase gradually when cultured under hypoxic conditions at 4–8 h, but then suddenly decrease at 24 h, when exactly is the largest increase in osteoclasts, suggesting that hypoxia-stimulated osteoclastogenesis may be not through increasing osteoclast viability. In summary, autophagy is a key regulator in hypoxia-induced osteoclastogenesis, highlighting that prolonged sustained force should not be applied during orthodontic clinical treatment. Further studies are needed to uncover the precise mechanisms of autophagy regulating osteoclastogenesis during OTM.
Autophagy in OTM-related CFCF is a major kind of orthodontic force and determines the speed of OTM. Numerous articles have explored the roles of CF in OTM; however, some conflicting results have emerged. Therefore, there is a great need to analyze the current studies so as to figure out the effects of autophagy in OTM and to guide clinical treatment. Specifically, we summarize the advances of autophagy in OTM-related cells (Fig. 5). Osteocytes and PDLCs are mechano-sensitive cells that respond to orthodontic forces primarily through numerous mechano-sensors present on the cell surface (Li et al. 2021b). Furthermore, PDLCs are mixed cells including fibroblasts (PDLFs), PDLSCs, osteoblasts, cementoblasts, osteoclasts, macrophages, and Malassez epithelial remnants. Therefore, we will present the roles of autophagy according to cell type in this part. However, due to the lack of research on Malassez epithelial remnants, they will not be discussed.
CF activates osteocyte autophagyOsteocytes are the primary sensor of mechanical signals and their autophagy is a research focus. Osteocyte autophagy is activated under CF in a murine OTM model, and CF and Rapa not only induce autophagy but also increase the secretion of Receptor activator of nuclear factor-kappa B ligand (RANKL) in osteocyte-like cell line MLO-Y4 cells by 3-fold and 4-fold, respectively (Li et al. 2020). Further, CF induces autophagy by reducing the expression of Rho-associated kinase (ROCK) and phosphorylation of protein kinase B (p-AKT) in a time-dependent manner while simultaneously increasing transcription factor E3 (TFE3). Consistently, Gao et al. report that autophagy is increased after exposure to cyclic CF in MLO-Y4 cells (Gao et al. 2023). Besides, adenosine triphosphate (ATP) metabolism, OCN, and cell survival are also increased in osteocytes via the mTOR complex 2 (MTORC2) activation, which is supported by another research indicating that MTORC2 activation protects cell survival via AKT signaling after autophagy induction (Wen et al. 2019). It seems confusing that MTORC2 has a potential role in inhibiting autophagy whereas autophagy activates MTORC2. It’s worth noting that MTORC2 and its downstream target protein kinase Ypk1 promote autophagy by negatively regulating Ca2+/calmodulin-dependent phosphatase and calcineurin during amino acid starvation (Vlahakis et al. 2014). Moreover, persistent MTORC1 inhibition can downregulate negative feedback loops on insulin-receptor substrate (IRS)-MTORC2-AKT to activate MTORC2 under starvation conditions (Bernard et al. 2014).
So, CF activates osteocyte autophagy. Additionally, both of the articles present the upregulation of LC3-II isoform and the degradation of Sequestosome 1/p62 (SQSTM1/p62, p62), collectively confirming an increased autophagic flux. Notably, they further investigate the effect of CF-stimulated osteocyte autophagy on osteoclastogenesis, which is in line with the reality of cellular interactions in vivo. Specific roles and related mechanisms will be introduced in later subgroups. Inadequate blood supply makes osteocytes survive in a hypoxic and nutrient-poor environment. Yet, osteocytes live for exceedingly prolonged lifespans. Therefore, further exploration is needed to assess how osteocytes adapt to their harsh surroundings, which will drive the development of OTM mechanisms.
CF induces autophagy in PDLCsPDLCs are another class of mechanical signal sensing and delivery cells that attract the attention of orthodontists and researchers. In 2019, Chen et al. reported that autophagy is increased in PDL tissues on the compression side during OTM, whereas human PDLC autophagy is initially increased but then decreased over time during CF application (Chen et al. 2019). 1.5 g/cm2 CF inhibits autophagy-related markers after 12 h and 4.5 g/cm2 CF for 6 h inhibits autophagy, providing a theoretical basis for the clinical application of forces. In general, autophagy response to stress can be divided into two phases: the first phase is a rapid increase in autophagic flux within minutes or hours after exposure to stress, and the second phase is a delayed and protracted stage (Pietrocola et al. 2013). In the study of Chen et al., PDLC autophagy is probably in the first phase. To better simulate the periodontal microenvironment, Zou et al. created a three‑dimensional (3D) PDLC culture in vitro using a collagen‑alginate composite hydrogel. They found that static compressive stress induces autophagy via integrin-linked kinase (ILK)/PI3K signaling pathway and upregulates ILK expression in a PI3K‑dependent manner (Zou et al. 2021). However, the western blot assay of Atg proteins is absent. That’s because total proteins can only be extracted after hydrogel dissolution, leading to protein degradation and affecting the result accuracy. Nevertheless, this article points out a 3D-PDLCs in vitro model and a promising study direction of PDLC autophagy under the OTM context.
In summary, autophagy is activated by CF in PDLCs. Nowadays, most OTM-related cellular models are two-dimensional (2D), but the 3D culture model does better to simulate cell growth in the extracellular matrix (ECM). Given the limitations of materials, it is crucial to explore hydrogels or better materials for 3D models that degrade quickly without side effects on cells. However, it is unfortunate that the above two studies directly use cells extracted from human PDL for research. It may be a trend to study one type of cell individually and to explore its interactions with other cells.
CF and inflammation activate autophagy in PDLFsPDLFs are the most numerous and functionally important cells in PDL that play a role in the transmission of mechanical stimulation and contribute to OTM (Li et al. 2018b). CF and inflammation activate autophagy in human PDLFs in a dose- and time-dependent manner via the mTOR pathway (Blawat et al. 2020). Interestingly, physiological pressure (2 g/cm2) has a cell-protective effect, whereas overload (8 g/cm2) and long-term Interleukin 1 beta (IL-1β) treatment increase cell death. Coincidentally, 2 g/cm2 CF or 0.1 ng/mL IL-1β treatment for a short time does not influence or even reduce autophagy, respectively. These data provide novel insights into autophagy regulation by pressure and inflammatory stress in PDLFs. However, direct evidence linking CF to cell death and autophagy is lacking, not to mention a specific regulatory mechanism. Thereafter, CF increases Interleukin 6 (IL-6) in PDLFs, which is further enhanced by 3-MA (Mayr et al. 2021). Consistently with in vitro studies, gene expressions of inflammatory factors IL-1, IL-6, and TNF-α are increased in periodontal tissues after applying a 0.35 N force along with 3-MA in a mouse OTM model (Chen and Hua 2021; Mo and Hua 2018). However, Mayr et al. didn’t find a significant IL-6 increase in a rat OTM with 25 g (Rath-Deschner et al. 2021), which may be attributed to the smaller force in a bigger animal.
Collectively speaking, PDLF autophagy is activated by both CF and inflammation. During orthodontic treatment, inflammation is critical for OTM, and inhibition of autophagy further upregulates inflammation that probably regulates OTM. Unfortunately, there is a significant discrepancy between the number of studies on PDLFs in OTM and their number in periodontal tissues. More research is urgently needed to explore the roles of autophagy in PDLFs and how they transmit mechanical signals and affect OTM.
Regulating PDLSC autophagy affects OTMPDLSCs respond to mechanical force and then contribute to the inflammatory response and alveolar bone remodeling during OTM (Huang et al. 2018). CF induces autophagy in a force-dependent and time-dependent manner in PDLSCs, and autophagic flux is upregulated under 1.5 g/cm2 CF stimuli within 6 h (Jiang et al. 2021). Besides, 1.0 and 1.5 g/cm2 CF triggered the highest ratio of LC3-II/I rather than 2.5 g/cm2, supporting that autophagy is sensitive to light CF in PDLCs. Importantly, the increased LC3 is attenuated by 3-MA injection in the periodontal tissues in an SD rat OTM model. Further, CF increases autophagy and strongly upregulates the expression of long non-coding RNA (lncRNA) FER1L4 in PDLSCs and in a murine OTM model (Huang et al. 2021b). Moreover, overexpression of FER1L4 increases the formation of autophagosomes and autolysosomes in PDLSCs, while knocking down FER1L4 reverses CF-induced autophagy. Mechanistically, FER1L4 inhibits p-AKT and increases the nuclear translocation of Forkhead box O3 (FOXO3), thereby upregulating autophagy under CF. However, the precise mechanism of FER1L4 inhibiting p-AKT needs further research. Additionally, lncRNAs may regulate ATG genes at the transcriptional, post-transcriptional, and translational levels, so other molecules may also be involved in the complicated process. Therefore, it expands our view that lncRNAs can be potential therapeutic targets for regulating OTM.
It’s worth noting that 2.0 g/cm2 CF lasting for 12 h has no significant influence on PDLSC apoptosis, but the apoptotic cells are significantly enhanced when autophagy is inhibited in the force group (Huang et al. 2021b). However, another group uses 2.0 g/cm2 CF to treat PDLSCs for 24 h and claims that CF significantly increases apoptosis and autophagy (Li et al. 2022a). To explain these differences, we need to know that autophagy is activated to remove damaged proteins and organelles and oppose apoptotic responses under CF, but if it fails, cell apoptosis and tissue damage will occur (Teng et al. 2011). Therefore, the apoptotic cells may increase when the autophagic flux is blocked by CQ in the former article. The increase in apoptotic cells in the latter one is likely caused by the prolonged CF stimulation time, which also suggests orthodontists use light and intermittent forces in the clinic.
Li et al. also demonstrate that the application of Apocynin attenuates long-term CF-induced apoptosis by regulating BECN1-mediated autophagy in PDLSCs, which provides an alternative approach to improve orthodontic treatment outcomes. However, there are several limitations to the study. Firstly, it requires a deeper study of the Apocynin-related signaling pathways in molecular cross-talk between autophagy and apoptosis during orthodontic treatment. Further, the specific regulation of the interaction between B cell leukemia/lymphoma 2 (Bcl2) and BECN1 with Apocynin during orthodontic treatment has not been fully identified. Most importantly, adding animal experiments to validate the effects of Apocynin is also a possible improvement.
In a word, the above articles clarify that autophagy activation serves as a protective mechanism under CF in PDLSCs in vivo and in vitro and contributes to bone remodeling and OTM. Nevertheless, the OTM mechanism is still mysterious, and PDLSCs with stem cell properties may play a significant role in OTM. Indeed, regulating autophagy, such as lncRNAs FER1L4 and Apocynin, affects OTM. In addition, apoptosis and autophagy are closely related, and how to obtain a balance between them to maximize orthodontic patient benefits will also be an interesting topic. In the future, RNA sequencing (RNA-seq) and gene editing technologies are urgently needed to explore more targets to optimize orthodontic treatments.
Supernatants from autophagy-activated cells promotes osteogenesisOsteoblasts can secrete and mineralize bone matrix. Since the first report of autophagy increase in osteoblast differentiation, accumulating evidence has demonstrated that suppression of autophagy reduces osteoblast mineralization and disrupts the balance between osteoblasts and osteoclasts (Wang et al. 2023). In osteoblasts, autophagy could be used as a vehicle to secrete apatite crystals, and a deficiency in autophagy leads to bone loss (Nollet et al. 2014). Supernatant collected from 3-MA-induced PDLFs decreases osteoprotegerin (OPG) expression in primary human alveolar osteoblasts (Mayr et al. 2021), highlighting the cross-talk between fibroblasts and osteoblasts and partly explains the in vivo findings that inhibition of autophagy in OTM decreases bone mineral density (Chen and Hua 2021). Similarly, the supernatant of MLO-Y4 cells under cyclic CF promotes osteoblast differentiation of murine osteoblasts MC3T3-E1 cells but inhibits osteogenesis by siATG7 in osteocytes (Gao et al. 2023). It may result from the increase of fibroblast growth factor 23 (FGF23) the decrease of sclerostin (SOST) in the supernatant, as the secretion of FGF23 (a homeostatic regulator within mineralization and phosphate) facilitates the development and formation of osteoblasts (Xu et al. 2022a) and the decrease of SOST (a bone morphogenetic protein (BMP) antagonist) due to cyclic CF-indued autophagy may enhance osteoblast differentiation (Chen et al. 2020b). However, the authors do not present direct evidence to prove the possible mechanism.
In addition, Jacox et al. observed no autophagic activity in osteoblasts in the Green Fluorescent Protein-LC3 (GFP-LC3) reporter mouse OTM model (Jacox et al. 2022). One possibility is that the alveolar bone is remodeled throughout life, and autophagy is necessary for osteogenesis, so autophagy doesn’t change much after the addition of force. Furthermore, given the complicated cell kinds in vivo, further studies are needed, such as a more extensive screen of cell-specific labels. More importantly, since osteoblasts are mechanically sensitive cells, more attention should be focused on the roles of CF in autophagy, as well as the underlying mechanisms. To date, the roles of CF in osteoblast autophagy during OTM remain understudied, but it is well worth exploring.
Intervening autophagy modulates CF-inhibited cementoblast autophagyDuring OTM, cementoblast proliferation and differentiation are modulated by orthodontic forces, which are closely correlated with OIIRR. Different from other PDLCs, cementoblast autophagy is suppressed under CF. Further, CQ reduces cellular migration, and Rapa partially relieves CF-inhibited cementoblast migration, which is partially dependent on matrix metalloproteinases (MMP9 and MMP13) (Yang et al. 2021). The different autophagic response to CF in cementoblasts and other PDLCs could be due to PDLCs adapting to CF more rapidly whereas cementoblasts exhibit slower adaptation, which also makes sense that it is primarily OTM caused by alveolar bone resorption rather than OIIRR of root apical resorption. In addition, 1.5 g/cm2 CF maintained for 12 h may be the optimal magnitude and treatment time in the immortalized murine cementoblast cell line OCCM-30 cells, whereas apoptosis is increased significantly after 24 h of CF and the ratio of apoptotic cells at 2.0 g/cm2 CF is significantly higher than that of the control group. The findings provide new insights into the role of autophagy in the biological behaviors of cementoblasts under CF; however, additional research is necessary to explore a protective role of autophagy in cementoblast migration in vivo and a potential therapeutic strategy for reducing OIIRR.
Additionally, CF impedes autophagy and downregulates mineralization-related markers, but strongly enhances lncRNA p21 expression in OCCM-30 cells (Liu et al. 2022). Furthermore, overexpression of lncRNA p21 downregulates autophagy and mineralization, while knockdown of lncRNA p21 reverses the effects. Importantly, 3-MA abolishes the lncRNA p21 knockdown-promoted mineralization, and Rapa rescues the mineralization inhibited by lncRNA p21 overexpression. Mechanistically, lncRNA p21 directly binds with FOXO3 and blocks autophagy. It should be mentioned that lentiviral inhibition of lncRNA p21 rescues impaired cementoblastic differentiation and effectively attenuates OIIRR. However, local injection of lentivirus could lead to off-target inhibition of ncRNAs in resident PDLCs, which may be a reason for inhibition of OTM by lncRNA p21 knockdown. Besides, Liu et al. report that knockdown of FOXO3 relieves the autophagy induced by lncRNA p21 deficiency. However, Lin et al. show that the downregulation of FOXO3 enhances the transcriptional activity of ATG genes and triggers the autophagic signaling pathway in liver cancer (Lin et al. 2020). LncRNA FER1L4 increases the nuclear translocation of FOXO3 thus activating autophagy in PDLSCs (Huang et al. 2021b). The above researches indicate the complex roles of FOXO3 in multiple forms of autophagy in different tissues and physiological conditions.
Similarly, CF inhibits cementoblast autophagy and mineralization, and autophagy activation markedly reverses cementoblast mineralization and prevents cementum damage in mice during OTM (Yang et al. 2023). As a mediator of autophagy and mineralization, Periostin silencing suppresses Wnt signaling by modulating the stability of β-catenin, thereby inhibiting mineralization. These data provide evidence that autophagy is indispensable to the restoration of CF-suppressed cementoblast mineralization in vitro and in vivo, and identify a new therapeutic target (Periostin) for cementum mineralization and periodontal tissue regeneration. Nevertheless, the application of CF on murine cementoblasts cannot exactly mimic the biological conditions on the compression side during OTM. Notably, a previous study has indicated that mechanical force regulates the osteogenic differentiation of PDLCs, where they load a frequency of 0.23 Hz at a 1.5 g/cm2 intermittent CF (Manokawinchoke et al. 2019). While Yang et al. demonstrate that 1.5 g/cm2 CF for 12 h suppresses cementoblast mineralization. These two studies apply different forms of force to various types of cells. However, it is still worth discussing, after all, they get opposite results of the effect of CF on cell mineralization. A possible reason is the distinctive roles of CF on cell autophagy, but further studies are needed to precisely locate the effects of autophagy in various cells and explore the underlying mechanisms.
In conclusion, cementoblast autophagy is suppressed under CF and they support that lncRNA p21 and periostin may serve as targets to inhibit autophagy and regulate cementogenesis. Cementoblasts contribute to the repair of cementum and restore periodontal function. Investigations into how CF impacts cementoblast mineralization are important for the development of therapeutics to repair OIIRR and achieve healthy periodontal function during OTM. We should make good use of high-throughput RNA-seq, as it is a simple and efficient technical means that helps us screen for therapeutic targets and identify potential pathways. Exploring more underlying mechanisms is crucial to provide theoretical guidance to the clinic.
Activating osteoclast autophagy accelerates OTMOsteoclasts are key cells for bone resorption and creating space for OTM. Cytokine (CF) functions in osteoclast precursor cell macrophages and is essential for maintaining tissue homeostasis. Previous studies have reported that CF promotes osteoclast differentiation and activation (Changkhaokham et al. 2022). Furthermore, CF increases autophagy and the RANKL/OPG ratio, and 3-MA further enhances the RANKL/OPG ratio and osteoclasts, so the authors speculate that autophagy might negatively regulate osteoclastogenesis (Chen et al. 2019). It is probably a shortcoming that osteoclasts are not co-cultured with PDLCs or treated with supernatants collected from PDLCs. Interestingly, activating autophagy by Rapa reduces the decline of bone density, downregulates osteoclasts and the RANKL/OPG ratio, and inhibits inflammation during OTM in vivo (Chen and Hua 2021; Chen et al. 2019). Differently, Li et al. report that CF-promoted osteocyte autophagy increases RANKL secretion through TFE3-related signaling, thereby enhancing osteoclastogenesis (Li et al. 2020). These opposite roles in osteoclastogenesis indicate that PDLCs and osteocytes may play different roles in tissue remodeling during OTM. Besides, due to the technical limitations of working with a minuscule mouse molar, it is difficult to separately analyze compression and tension tissues in PDL, let alone specific cell components.
However, the two teams report controversial results, even though they both study the roles of osteocyte autophagy in osteoclastogenesis. Gao et al. culture a mouse osteoclast precursor cell line RAW 264.7 cells in supernatants collected in cyclic CF-induced osteocytes and observe that inhibiting osteocyte autophagy by siATG7 attenuates the downregulation of osteoclastogenesis by MTORC2 activation (Gao et al. 2023), which is opposite with Li et al. The possible reason might be the different types of CF and different loading durations, as Li et al. apply a 0.5 g/cm2 CF for 1, 3, and 6 h whereas Gao et al. use a cyclic CF at a frequency of 2 Hz with 2000 µε for 10 min, 30 min, 1 h, 3 h, and 6 h. Further studies are needed to identify the effect of direct interplay with osteocytes and other cells, and co-culture is an excellent example. Surely, osteoclasts can also be treated by exosomes from osteocytes. As the cytokine secretion of RANKL, OPG, or macrophage-stimulating factor (M-CSF) from osteocytes, the autocrine and paracrine function may be another research point.
Regulating autophagy may change force-induced autophagy on osteoclast activity. As previously mentioned, blockage of autophagy by 3-MA increases the expression of osteoclast-related markers, decreases bone density, and promotes tooth movement in the murine OTM model (Chen et al. 2019). Supportively, intraperitoneal injection of 3-MA rescues the reduced osteoclastogenesis and promotes OTM in a nucleotide-binding domain (NBD), leucine-rich repeat (LRR), and pyrin domain (PYD)-containing protein 3 deficient (NLRP3−/−) mice (Han et al.
Comments (0)