
Remember me





We measured serum T4 to assess efficacy of the mouse diets to induce the hypo- or hyperthyroid state. The hypothyroid diet resulted in a greater than 4-fold decrease in T4 levels in females (FPTU: p = 0.014, Cohen’s d = 3.4, 95% CI [0.6363, 6.082]) and males (MPTU: p = 0.017, Cohen’s d = 6.66, 95% CI [0.4997, 5.594]). The TH diet resulted in a greater than 3-fold increase in T4 levels in females (FTH: p < 0.0001, Cohen’s d = 4.6, 95% CI [−11.95, − 6.501]) and males (MTH: p < 0.0001, Cohen’s d = 23.40, 95% CI [−13.63, − 8.189]) (Fig. 2). Thus, our dietary interventions had their intended effects on thyroid hormone levels.
Fig. 2
Serum T4 concentrations in aged male (M) and female (F) mice. Euthyroid (CON), hypothyroid (PTU) and hyperthyroid (TH). Data presented with bars for means and one standard deviation. Statistics – one-way ANOVA – correct for multiple comparison- test: Šidák post hoc analysis was performed (n = 3 animals/sex/treatment).
Sex based differences in morphometric and echocardiographic responses to altered thyroid hormone levelsWe performed serial echocardiography to assess changes in left ventricular function compared to baseline values in response to diet treatments (Table 1). The FTH hearts had significantly improved left ventricular systolic function as measured by increased ejection fraction (p = 0.0003, Cohen’s d = 5.24, 95% CI [−34.23, − 11.26]) and fractional shortening (p = < 0.0001, Cohen’s d = 6.2, 95% CI [−22.44, − 8.34]). We observed no significant changes in left ventricular systolic function between pre- and post-treatment for any other groups.
Table 1 Echocardiographic and morphometric analysis of the heart after four weeks on prescribed dietThe FTH hearts had a significantly higher heart rate (p = 0. 012, Cohen’s d = 2.4, 95% CI [−225.7, − 25]) during the final echocardiogram compared to baseline pre-treatment. Both hypothyroid (MPTU and FPTU) and MTH groups had no differences in functional parameters between pre- and post-treatment.
We next evaluated cardiac hypertrophy by measuring heart weight normalized to tibia length (HW/LT, Table 1). MPTU showed a 23% lower HW/TL ratio compared to MCON (p = 0.06, Cohen’s d = 1.76, 95% CI [−0.04, 3.02] suggesting a shift toward cardiac atrophy although it did not reach significance. We found no difference in FPTU compared to FCON. TH increased HW/TL ratio for both male and female hearts by 26% (p = 0.001, Cohen’s d = 6.3, 94%CI[[−4.22, − 1.16]) and 51% (p = < 0.0001, Cohen’s d = 12.3, 95% CI [−5.51, − 2.45]) respectively compared to their controls (Table 1).
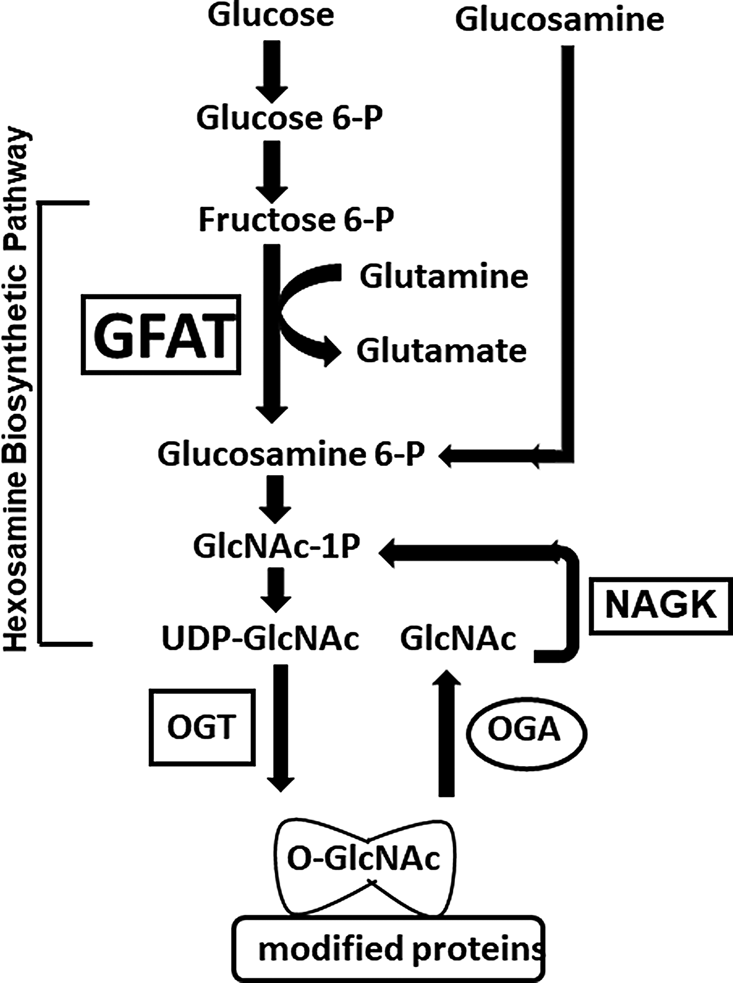
Predicted thyroid hormone receptor element (TRE) binding sitesTo assess the potential of TH regulating O-GlcNAc, we did a search of the JASPAR 2020_NR database for TREs in the O-GlcNAc regulatory proteins: OGT, OGA, GFAT1 and GFAT 2 using the PSCAN software (http://www.beaconlab.it/pscan) [13]. The search identified potential TRE sites upstream of the genes; however, only OGT and OGA had significant p-values (Table 2).
Table 2 Predicted thyroid hormone receptor binding sitesSex-based differences in global cardiac O-GlcNAc levels and its regulatory proteinsWe used western blots to assess total protein O-GlcNAc levels, as well as the key O-GlcNAc regulatory proteins - GFAT1, GFAT2, OGA, OGT and NAGK.
O-GlcNAc levels (Fig. 3 and 4). We measured total O-GlcNAc across the molecular weights spanning 100 kDa to 25 kDa in the heart (Fig. 3). Both FPTU (p = 0.003, Cohen’s d = 15.3, 95% CI [−0.9, − 0.20]) and FTH (p = 0.0002, Cohen’s d = 10.5, 95% CI [−1.1, − 0.39]) had significantly greater global O-GlcNAc levels compared to FCON. MTH also had higher global O-GlcNAc versus MCON (p = 0.0003, Cohen’s d = 3, 95% CI [−1.1, − 0.35]). Differing from the females, however, MPTU had no effect on global O-GlcNAc levels in male hearts.
Fig. 3
Total heart protein O-GlcNAcylation levels in males (M) and females (F) comparing euthyroid (CON), hypothyroid (PTU) and hyperthyroid (TH). All values are arbitrary units normalized to total protein levels. Data presented with bars for means and standard deviation. Statistics – one-way ANOVA – correct for multiple comparison- test: Šidák post hoc analysis was performed (n = 3 animals/sex/treatment). Image was cropped to obtain the PTU/CON/TH arrangement. Full-length uncropped blots are presented in Supplementary Fig. 1
Fig. 4
Total liver protein O-GlcNAcylation levels in males (M) and females (F) comparing euthyroid (CON), hypothyroid (PTU) and hyperthyroid (TH). All values are arbitrary units normalized to total protein levels. Data presented with bars for means and one standard deviation. Statistics – one-way ANOVA – correct for multiple comparison- test: Šidák post hoc analysis was performed (n = 3 animals/sex/treatment). Full-length uncropped blots are presented in Supplementary Fig. 1
We also wanted to assess whether O-GlcNAc changes were specific to the heart. Therefore, we measured liver O-GlcNAc levels (Fig. 4). The MTH livers, like the MTH hearts, showed significantly increased O-GlcNAc levels (p = 0.001, Cohen’s d = 6.3, 95% CI [−1.22, − 0.33]). Similarly, the MPTU livers, like the MPTU hearts were unchanged under the hypothyroid conditions. The female liver showed no significant change in O-GlcNAc levels due TH status which represents an important difference from female hearts.
We subsequently assessed candidate mechanisms whereby TH alters O-GlcNAc levels in the heart.
GFAT protein levels in the heart (Fig. 5 and 6). GFAT is the rate limiting step in the HBP and has two isoforms, GFAT1 and GFAT2. GFAT1 (p = 0.004, Cohen’s d = 3.4, 95% CI [−2.01, − 0.38]) and GFAT2 (p = 0.007, Cohen’s d = 6.1, 95% CI [−1.95, − 0.31]) levels increased in the FTH groups compared the FCON. GFAT1 and GFAT2 showed no significant difference for MPTU or MTH compared to MCON or FPTU compared to FCON.
Fig. 5
Heart protein immunoblots for GFAT 1 in males (M) and females (F) comparing euthyroid (CON), hypothyroid (PTU) and hyperthyroid (TH). Immunoblots are shown for GFAT1 protein levels normalized to total protein stain. Data presented with bars for means and one standard deviation. Statistics – one-way ANOVA – correct for multiple comparison- test: Šidák post hoc analysis was performed (n = 3 animals/sex/treatment). The asterisk – unknown band. Full-length uncropped blots are presented in Supplementary Fig. 1
Fig. 6
Heart protein immunoblots for GFAT2 in males (M) and females (F) comparing euthyroid (CON), hypothyroid (PTU) and hyperthyroid (TH). Immunoblots are shown for GFAT2 protein levels normalized to total protein stain. Data presented with bars for means and one standard deviation. Statistics – one-way ANOVA – correct for multiple comparison- test: Šidák post hoc analysis was performed (n = 3 animals/sex/treatment). Image was cropped to obtain the PTU/CON/TH arrangement. Full-length uncropped blots are presented in Supplementary Fig. 1
OGT protein levels in the heart (Fig. 7). OGT attaches the O-GlcNAc moiety to the target proteins. FPTU (p = < 0.0001, Cohen’s d = 3.9, 95% CI [−2, − 0.87]) and FTH (p = 0.0007, Cohen’s d = 7, 95% CI [−1.67, − 0.49]) both had increased OGT protein levels compared to FCON. Males showed no change in OGT expression in response to TH status.
Fig. 7
Heart protein immunoblots for OGT in males (M) and females (F) comparing euthyroid (CON), hypothyroid (PTU) and hyperthyroid (TH). Immunoblots are shown for total protein OGT levels normalized to total protein stain. Data presented with bars for means and one standard deviation. Statistics – one-way ANOVA – correct for multiple comparison- test: Šidák post hoc analysis was performed (n = 3 animals/sex/treatment). Full-length uncropped blots are presented in Supplementary Fig. 1
OGA protein levels in the heart (Fig. 8) OGA removes the O-GlcNAc moiety from proteins. The MTH group (p = 0.03, Cohen’s d = 8.2, 95% CI [0.03, 0.75]) was the only group to show a difference compared to their control group.
Fig. 8
Heart protein immunoblots for OGA in males (M) and females (F) comparing euthyroid (CON), hypothyroid (PTU) and hyperthyroid (TH). Immunoblots are shown for total protein OGA levels normalized to total protein stain Data presented with bars for means and one standard deviation. Statistics – one-way ANOVA – correct for multiple comparison- test: Šidák post hoc analysis was performed (n = 3 animals/sex/treatment). Image was cropped to obtain the PTU/CON/TH arrangement. Full-length uncropped blots are presented in Supplementary Fig. 1
NAGK protein levels in the heart (Fig. 9). N-acetylglucosamine kinase recycles GlcNAc back into the HBP for reuse in protein O-GlcNAcylation. Both FPTU (p = < 0.0001, Cohen’s d = 35, 95% CI [−1.48, − 0.84]) and the MTH (p = < 0.0001, Cohen’s d = 6, 95% CI [−1.4, − 0.74]) groups had increased NAGK levels compared to their control group. We found no changes for the other groups.
Fig. 9
Heart protein immunoblots for NAGK in males (M) and females (F) comparing euthyroid (CON), hypothyroid (PTU) and hyperthyroid (TH). Immunoblots are shown for total protein NAGK levels normalized to total protein stain. Data presented with bars for means and one standard deviation. Statistics – one-way ANOVA – correct for multiple comparison- test: Šidák post hoc analysis was performed (n = 3 animals/sex/treatment). Full-length uncropped blots are presented in Supplementary Fig. 1
Sex-based differences in O-GlcNAc and its regulatory proteins transcripts in the heartGiven that TH can act through its receptors to alter gene transcription we assessed changes in cardiac transcript levels of the key O-GlcNAc regulatory proteins - GFAT1, GFAT2, OGA, OGT and NAGK (Table 3).
Table 3 Fold Change (2^ΔΔCt) in gene expression by qPCR of cardiac O-GlcNAc regulatory proteinsGFAT transcript levels in the heart. We found higher GFAT1 transcript levels in MPTU compared to the MCON group (p = 0.005, Cohen’s d = 3.5, 95% CI [−0.53, − 0.13]) and lower GFAT1 transcript levels in MTH (p = 0.01, Cohen’s d = 2.8, 95% CI [0.07, 0.46]), but no differences for the female groups. GFAT2 transcript levels were unchanged for all groups in both sexes.
OGT transcript levels in the heart. MPTU (p = 0.03, Cohen’s d = 2.3, 95% CI [−0.67, − 0.44]) transcript levels increased compared to MCON group. There were no differences for the other groups.
OGA transcript levels in the heart. MPTU (p = 0.002, Cohen’s d = 4.1, 95% CI [−0.55, − 0.19]) transcripts showed increased levels compared to the MCON group, whereas the FPTU group was unchanged to the FCON group. Both the MTH (p = 0.002, Cohen’s d = 4.2, 95% CI [0.19, 0.55]) and FTH (p = 0.04, Cohen’s d = 2.5, 95% CI [0.016, 0.59]) showed decreased OGA transcript levels compared to their respective group.
NAGK transcript levels in the heart. MPTU (p = < 0.0001, Cohen’s d = 7.2, 95% CI [−1.16, − 0.66]) transcripts showed increased levels compared to the MCON group, whereas the FPTU group was unchanged to the FCON group. Both the MTH (p = 0.004, Cohen’s d = 4.2, 95% CI [0.18, 0.68]) and FTH (p = 0.03, Cohen’s d = 2.7, 95% CI [0.06, 0.72]) showed decreased NAGK transcript levels compared to their respective group.
Global protein O-GlcNAcylation patterns do not necessarily reflect changes on individual proteins in the heartWe wanted to determine how changes in global O-GlcNAc manifest at the level of a specific protein. We evaluated ACAA2 O-GlcNAcylation because we previously found that it is O-GlcNAcylated in the heart [21] and it also interacts with thyroid hormone receptor β1 [26]. To assess ACAA2 O-GlcNAc levels we performed two IPs targeting either the O-GlcNAc modification or the ACCA2 protein. To ensure our immunoprecipitation data was not from non-specific background activity, we used either a mouse IgG or rabbit IgG as a negative control (Fig. 10A and 10B inset) and found non-specific IgG antibody binding to be insignificant. Our results are shown in Fig. 10A and 10B. ACAA2 O-GlcNAcylation was unchanged in MPTU versus MCON regardless of immunoprecipitation antibody, which was expected since they had similar global O-GlcNAc levels. Matching the rise in global O-GlcNAc levels, O-GlcNAcylation of ACAA2 was greater in FPTU versus FCON, (RL2 IP: p = 0.002, Cohen’s d = 3, 95% CI [−3.5, − 0.8]) (ACAA2 IP: p = 0.002, Cohen’s d = 24, 95% CI [−0.59, − 0.42]). O-GlcNAc levels on ACAA2 for the hyperthyroid groups, however, were incongruent with the group’s high global O-GlcNAc levels. The FTH ACAA2 O-GlcNAc levels was unchanged compared to the FCON, whereas the MTH to MCON ACAA2 O-GlcNAC levels were decreased (ACAA2 bait: p = 0.01, Cohen’s d = 9.5, 95% CI [0.22, 0.60]). With the RL2 immunoprecipitation, MTH ACAA2 O-GlcNAc appeared decreased; however, the high standard deviation in the MCON groups negatively impacts the significance.
In total, we found O-GlcNAc protein specificity differs based upon stimulus (hypo- versus hyperthyroidism) and gender.
Fig. 10
Immunoblots for O-GlcNAc immunoprecipitated ACAA2 in males (M) and females (F) comparing euthyroid (CON), hypothyroid (PTU) and hyperthyroid (TH). (A) Immunoblots show ACAA2 expression in heart tissue immunoprecipitated with the O-GlcNAc RL2 antibody normalized to total ACAA2. Statistics – one-way ANOVA – correct for multiple comparison- test: Šidák post hoc analysis was performed (n = 3 animals/sex/treatment). Asterisks denotes a non-specific band. (B) Immunoblots of O-GlcNAc expression on immunoprecipitated ACAA2 protein. O-GlcNAc bands were normalized to input ACAA2. Statistics – Unpaired t test was performed (n = 2 animals/sex/treatment). Data presented with bars for means and one standard deviation. Full-length uncropped blots are presented in Supplementary Figure 1
Effect of O-GlcNAc levels on ACAA2 transcriptional activityTo assess the functional consequences of ACAA2 O-GlcNAcylation, we performed a luciferase reporter assay. Our lab has previously shown ACAA2 acts as a T3-dependent activator to the thyroid receptor β1 (TRβ1) protein [26], therefore, we investigated the effect of altering O-GlcNAc using either thiamet G, an OGA inhibitor to increase global O-GlcNAc levels, or OSMI-1, an OGT inhibitor to reduce O-GlcNAc levels. We used CV-1 cells due to the lack of endogenous thyroid receptor activity [27]. The OSMI-1 treated cells showed a significant decrease in T3 based ACAA2 activation of TRβ1 suggesting that lowering ACAA2 O-GlcNAcylation reduces TRβ1/ACAA2 transcriptional activity (Fig. 11).
Fig. 11
ACAA2/TRβ1 transcriptional assay. Inhibiting O-GlcNAc in transfected TH-treated CV-1 cells reduced ACAA2 activation of TRβ1 transcriptional activity. TRβ1 – thyroid receptor β1; ACAA2 – acetyl-coenzyme A acyltransferase; T3 – thyroid hormone; TG – thiamet G, OGA inhibitor; OSMI-1 – OGT inhibitor. Statistics – one-way ANOVA – correct for multiple comparison- test: Šidák post hoc analysis was performed (n = 4).
Comments (0)