
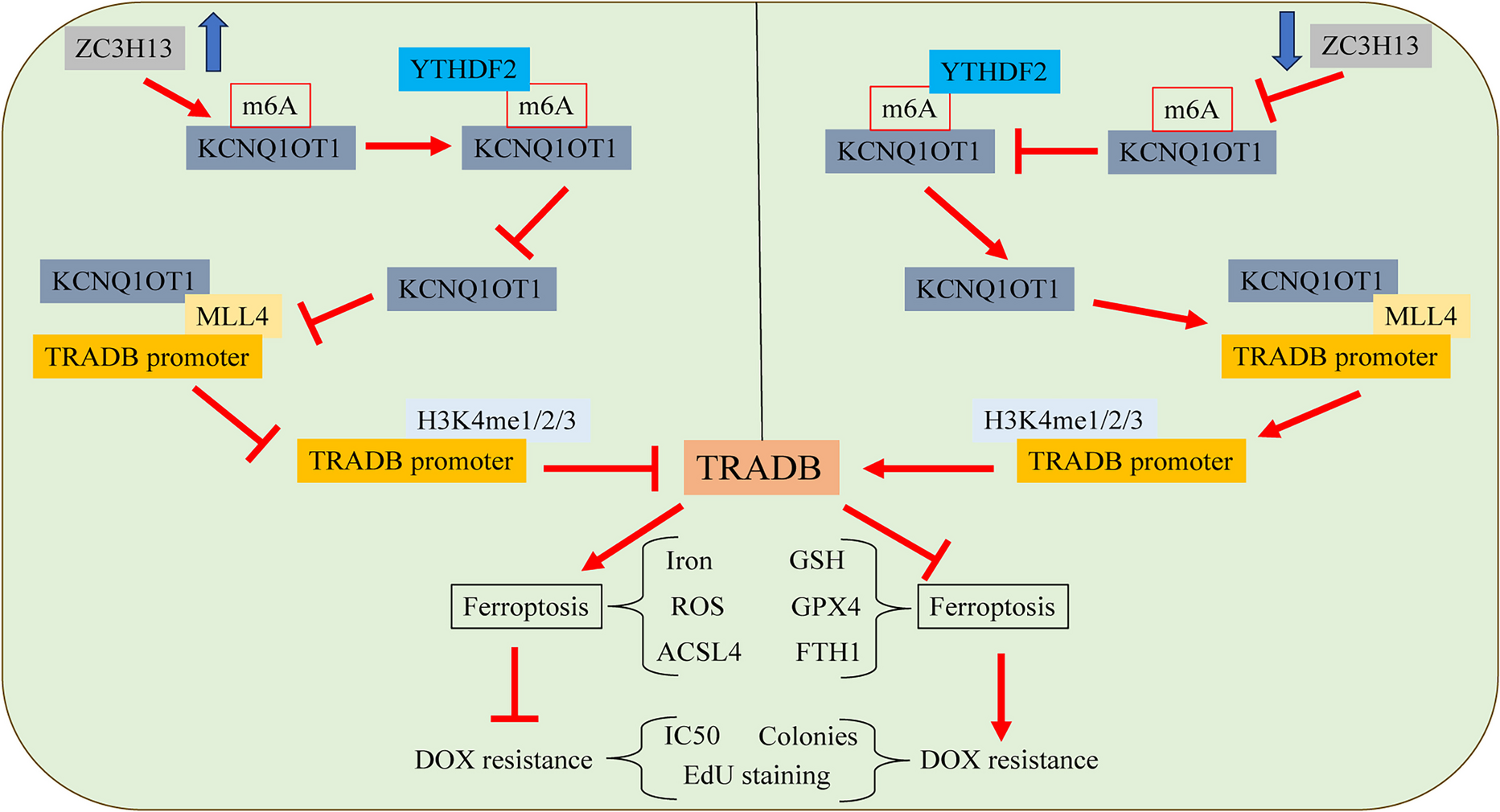
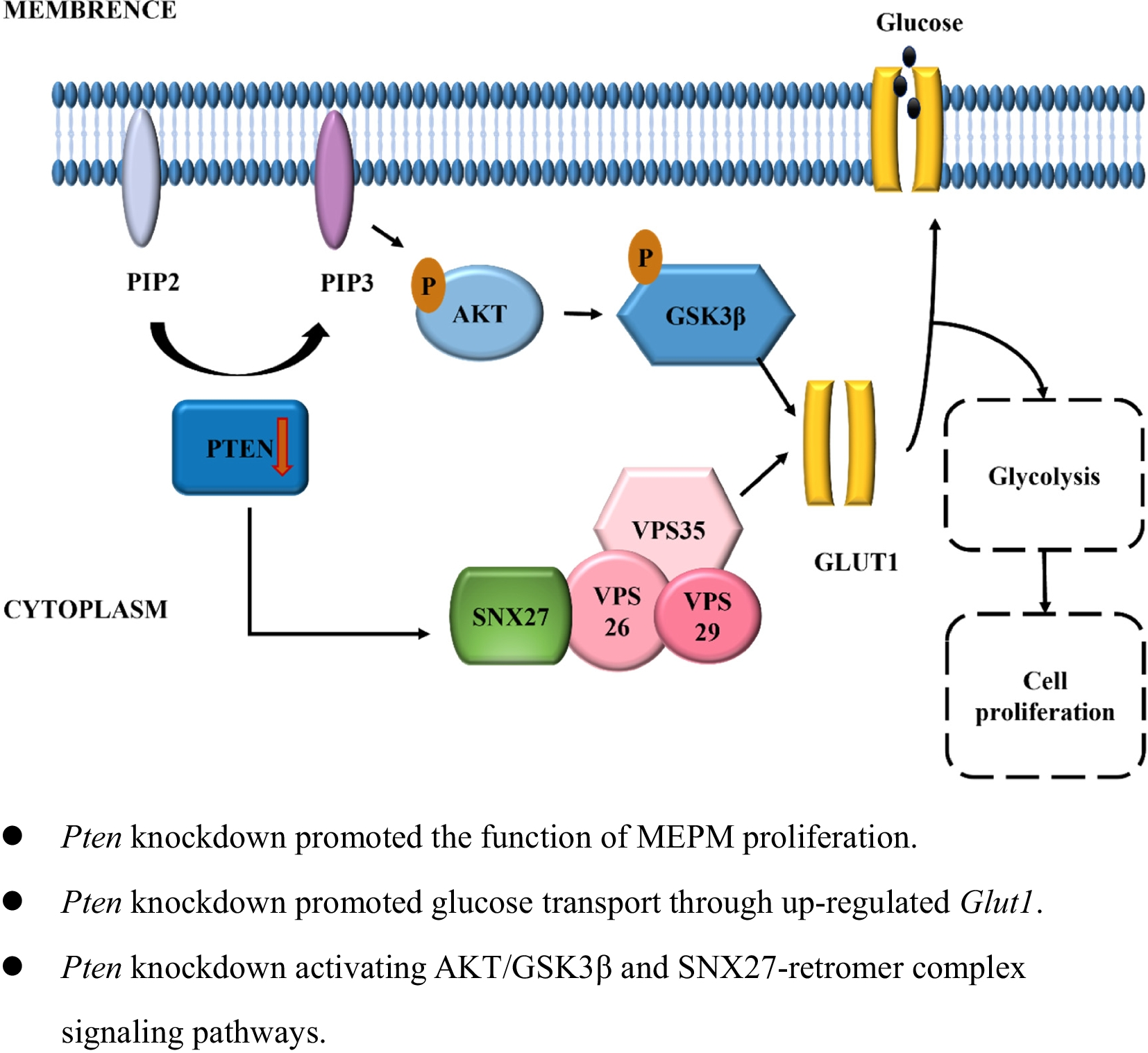
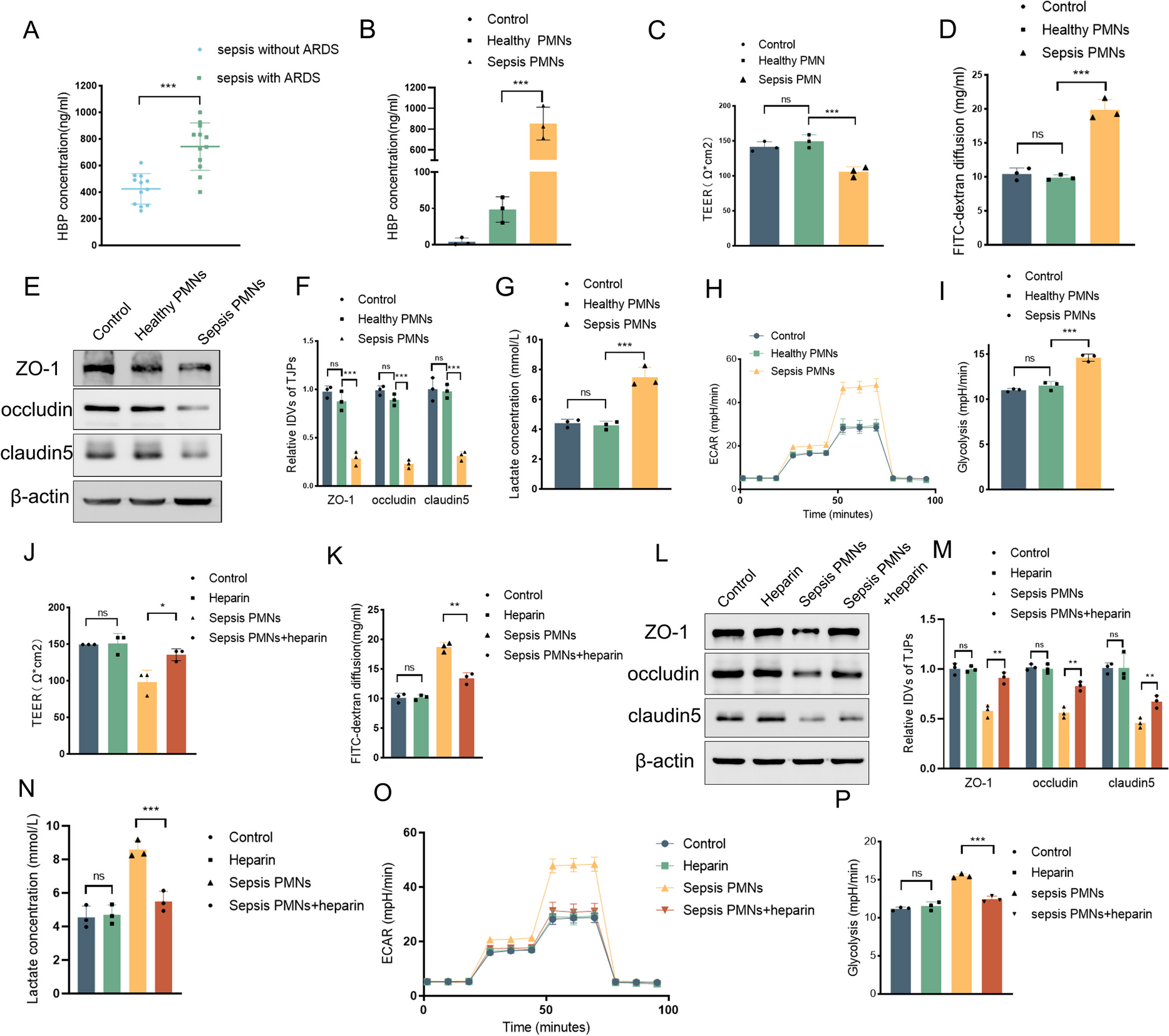
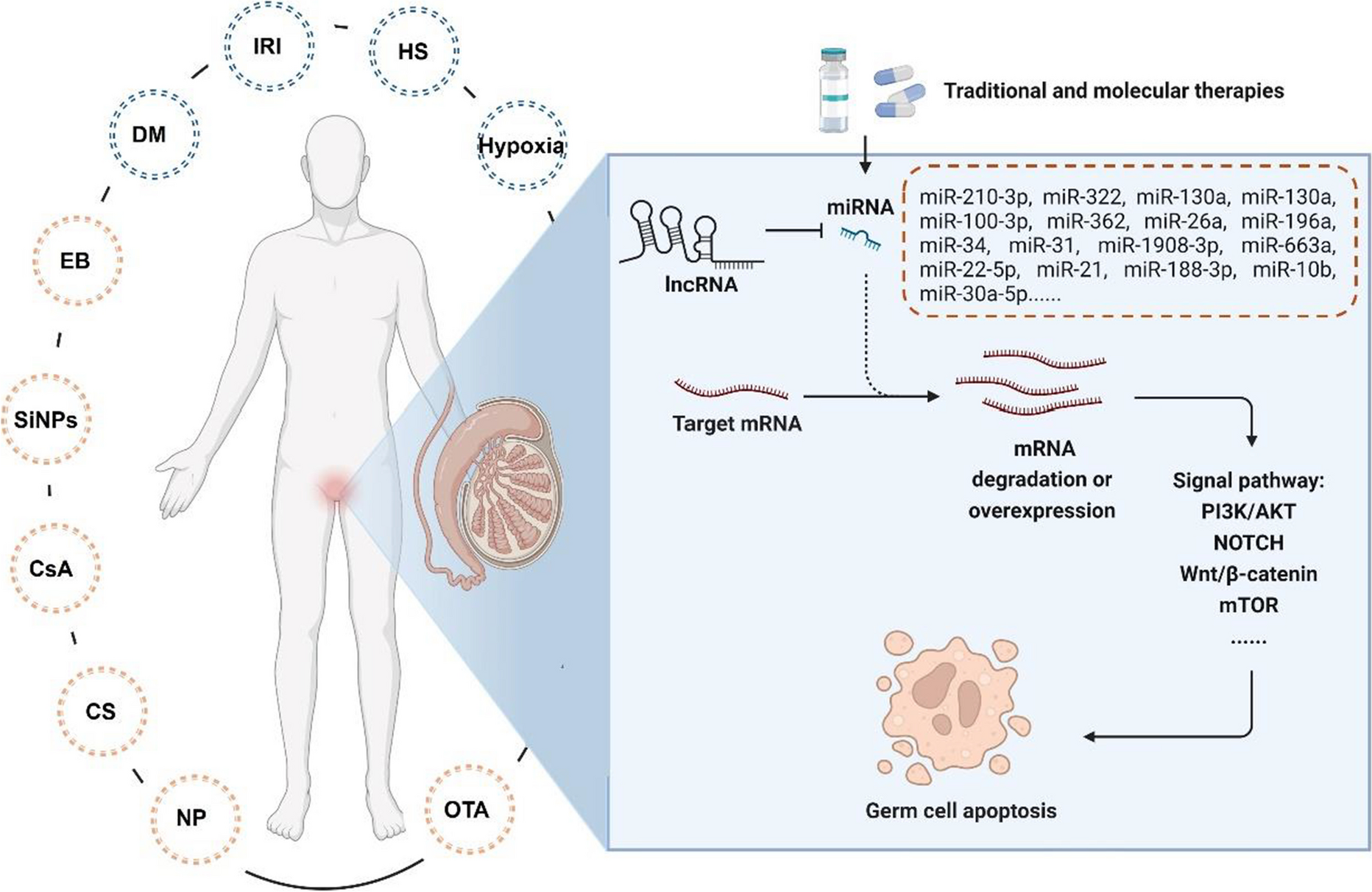
Remember me



To identify hub genes involved in the sorafenib resistance, we performed the WGCNA using the mRNA expression profiles of HCC patients in the GSE109211 dataset (Pinyol et al. 2019), which included a sorafenib treatment group (n = 67) and a placebo group (n = 73) (Fig. 1A and B). As a result, the brown module showed the strongest correlation with non-responders, with a Pearson correlation coefficient of 0.76 (Fig. 1C). Moreover, most genes in the brown module were upregulated in non-responders compared to responders (Fig. 1D). We then defined 35 hub genes with the module membership scores more than 0.85 and gene significance scores greater than 0.75 (Fig. 1E). To further determine the most important molecular players, we established a protein–protein interaction network using the 35 genes in STRING database, and identified 8 genes with the highest degree of connectivity in the network, including CUL1, SKP2, NBR1, NEDD8, PSMD4, PPP2R5C, VMP1, and PRMT1 (Fig. 1F). Intriguingly, CUL1 and SKP2 are key components of SCF complex (S: SKP2, C:CUL1 and F: F-box protein E3 ubiquitin ligases). CUL1 and SKP2 along with the genes NEDD8 for cullins neddylation formed a small network. In addition, NBR1, an autophagy receptor, was also included in the network.
Fig. 1
Identification of hub genes associated with sorafenib resistance in HCC. (A) The clustering dendrogram of sorafenib treatment responder and non-responder. (B) Color-represented clustering dendrogram of gene co-expression modules identified after using a merged dynamic tree cut. (C) Correlation between the sorafenib responsiveness and each module. (D) Heatmap of differentially expressed genes in the brown module. (E) Scatter diagram of genes in brown module. X-axis represents module membership, and Y-axis represents gene significance. (F) Identification of the most important proteins with the highest degree of connectivity within the protein–protein interaction network in STRING. (G) Enrichment analysis of differentially expressed genes between sorafenib non-responders and responders (left, upregulated; right, downregulated). (H) Box plots of CUL1, SKP2, and NEDD8 expression in GSE109211 cohort. (I) Expression of SCF E3 ubiquitin ligase complex components (CUL1, SKP2, and NEDD8) in sorafenib responders and non-responders in GSE109211 dataset. (J) Kaplan–Meier curves of HCC patients with high or low SKP2, CUL1, and NEDD8 expression in GSE109211 cohort
Beyond that, we conducted enrichment analysis based on the differentially expressed genes between sorafenib non-responders (n = 46) and responders (n = 21). The results showed that upregulated genes in sorafenib non-responders were enriched in protein neddylation, ubiquitination, post-translational protein modification, and autophagy (Fig. 1G), which further verified the reasonability of the above network. Coordinating the above results, it was expected that these genes (CUL1, SKP2, NBR1, and NEDD8) may play many pivotal roles in sorafenib resistance.
To further explore the significance of these genes in HCC development and progression, we assessed their expression and prognostic implications in GSE144269 and TCGA cohorts. The results showed that CUL1, SKP2, and NEDD8 were not only highly expressed in cancerous than normal tissues but also elevated in sorafenib non-responders than responders in GSE109211 dataset (Fig. 1H and I). The high expression of these genes in tumor tissues were strongly associated with poor outcomes (Fig. 1J). There was a similar trend in TCGA cohort (Supplementary Fig. 1). Notably, all these genes were highly expressed in cancerous than normal tissues (Supplementary Fig. 1A) and the high expression of these genes in tumor tissues were strongly associated with poor outcomes in TCGA dataset (Supplementary Fig. 1B). It also should be noted that an E3 NEDD8 ligase MDM2 is not only increased in cancerous than normal tissues (Supplementary Fig. 1A) but also elevated in sorafenib non-responders than responders (Fig. 1I). Therefore, it might facilitate to the neddylation modification. Indeed, as indicated by western blot assay, the knockdown of MDM2 decreased the CUL1 neddylation, but had no effect on the total protein level of CUL1 and SKP2 (Supplementary Fig. 1C).
Expression levels of hub genes in HCC patientsTo further investigate the expression of these genes, we collected the adjacent normal tissues and tumor tissues from 10 HCC patients who have been treated with or without sorafenib. We compared the gene expression in the normal tissues, primary tumor tissues (untreated with sorafenib) and tumor tissues (acquired drug resistance after sorafenib treatment). The PCR results showed no significant difference in expression of CUL1 and SKP2 between adjacent non-cancerous tissue and primary liver cancer tissue. However, CUL1 and SKP2 expression were higher in sorafenib-resistant tumors than in primary tumors (untreated with sorafenib) (Fig. 2A). As indicated by the western blot, CUL1 protein showed two bands with molecular weight 90KD and 85KD, respectively (Fig. 2B). The band of CUL1 protein with the molecular weight 90KD was significantly increased in the sorafenib-resistant tumor tissues compared to the primary tumor tissues, while no significant difference was observed in the band with molecular weight 85KD. The high molecular wight of CUL1 protein implies some protein modification. According to the evidence from the bioinformatics analysis, we tested the NEDD8-modified Cullin. NEDD8-Cullin and SKP2 were also significantly elevated in the sorafenib-resistant tumor tissues, compared to the primary tumor tissues. IHC assay showed that CUL1 was located in the cytoplasm in part of the cells, while, in other parts of cells, CUL1 was enriched in the cell nucleus (Fig. 2C). Based on the above results, it is conceived that post-translational modification such as neddylation in CUL1 was increased in sorafenib-resistant tumor.
Fig. 2
Expression of SCF components in sorafenib-resistant liver tumors. (A) qPCR analysis of CUL1 and SKP2 in adjacent normal tissues, primary tumor tissues without sorafenib treatment, and primary tumor tissues resistant to sorafenib. (B) Western blot of the SCF E3 ubiquitin ligase complex components (CUL1 and SKP2) and NEDD8-Cullin in adjacent normal tissues, primary tumor tissues without sorafenib treatment, and primary tumor tissues resistant to sorafenib. (C) Representative IHC images of CUL1 in HCC patients.*P < 0.05, **P < 0.01 and ***P < 0.001 (n = 10)
CUL1 is involved in the sorafenib resistance in HCCWe previously established sorafenib-resistant cell lines by exposing liver cancer cells (HepG2 and Huh7 parental cells) to sorafenib stimulation for 6 months at concentrations lower than their respective half maximal inhibitory concentration (IC50) values (Zhai et al. 2014). The resultant cell lines were then continuously cultured in the presence of sorafenib to maintain their drug-resistant characteristics. The CCK-8 assay showed that IC50 values of HepG2 parental cells (HepG2-P) and HepG2 sorafenib-resistant cells (HepG2-SR) were 4.33 μM and 12.84 μM, respectively (Fig. 3A). IC50 values of Huh7 parental cells (Huh7-P) and Huh7 sorafenib-resistant cells (Huh7-SR) were 3.98 μM and 12.05 μM, respectively. The resistant cells exhibited apparent advantages in cell viability (Fig. 3A), proliferation (Fig. 3B and Supplementary Fig. 2A), colony formation capacity (Fig. 3C and Supplementary Fig. 2B), and invasion (Fig. 3D) under higher dosages of sorafenib treatment, further demonstrating the successful establishment of resistant cells. As shown in western blot assay, NEDD8-Cullin and SKP2 were also significantly elevated in the HepG2-SR and Huh7-SR cells than in HepG2-P and Huh7-P cells, respectively (Fig. 3E and Supplementary Fig. 2C). We knocked CUL1 down in the HepG2-SR and Huh7-SR cells (Fig. 3F). The knockdown of CUL1 remarkably enhanced the suppressive effect of sorafenib on the cell viability, proliferation, colony formation capacity and invasion (Fig. 3G-J and Supplementary Fig. 2D, E). These results collectively suggest that CUL1 is involved in the sorafenib resistance in HCC.
Fig. 3
CUL1 is involved in the sorafenib resistance in HCC. (A) CCK-8 assay of parental cells (HepG2-P and Huh7-P) and sorafenib-resistant cells (HepG2-SR and Huh7-SR) in the presence of gradient sorafenib treatment. (B) EdU assay, (C) colony formation assay, and transwell assay (D) of HepG2-P, Huh7-P, HepG2-SR and Huh7-SR cells under 0, 5, and 10 μM sorafenib treatment. (E) Western blot of SCF components (CUL1 and SKP2), and NEDD8-Cullin in HepG2 and Huh7 parental cells and their corresponding sorafenib-resistant cells. (F) Western blot test of CUL1 after sorafenib treatment and CUL1 knockdown. (G) CCK-8 assay, (H) EdU assay, (I) colony formation assay, and transwell assay (J) of HepG2-P, Huh7-P, HepG2-SR and Huh7-SR cells after sorafenib treatment and CUL1 knockdown. *P < 0.05, **P < 0.01 and ***P < 0.001 (n = 3). Sor:sorafenib; KD: knockdown
CUL1 neddylation mediates p27 ubiquitination in sorafenib-resistant liver cancer cellsThe SCF complex is a subtype of Cullin-RING E3 ubiquitin ligase that includes CUL1 as its core Cullin protein. The E3 ligase facilitates the transfer of ubiquitin from E2 enzymes to target proteins, thus marking them for degradation by the proteasomes or lysosomes (Yang et al. 2021). The complete activation of ubiquitin ligases E3 ubiquitin ligases requires Cullin neddylation, a reversible modification by adding the ubiquitin-like protein NEDD8 to Cullins. Upon neddylation modification, ubiquitin ligases are activated to ubiquitinate specific substrates, such as various tumor suppressors (Enchev et al. 2015). p27 is a cyclin-dependent inhibitor that hindered the assembly and activity of cyclin-dependent kinases and cyclin complexes, thus inducing cell cycle arrest and inhibiting cell proliferation, finally causing cell apoptosis (Parazzi et al. 2015). However, p27 is usually degraded by SCF complex in cancer, leading to the uncontrolled proliferation of cancer.
Given the increased expression of SCF component proteins in sorafenib-resistant cells, it was hypothesized a decrease in p27 as a result of CUL1 neddylation-mediated ubiquitination and degeneration. To test this hypothesis, the resistant cells were treated with a neddylation inhibitor, MLN4924 (Zheng et al. 2021). Western blot results showed a significant decrease in NEDD8 and CUL1 expression, while MLN4924 treatment had little effect on SKP2 expression (Fig. 4A and Supplementary Fig. 3A). Notably, p27 expression was restored in resistant cells in a dose-dependent manner (Fig. 4A and Supplementary Fig. 3A). Moreover, the co-immunoprecipitation results showed the reduced amounts of p27-immunoprecipitated CUL1 and ubiquitin in the presence of MLN4924 (1 μM, Fig. 4B) or under the conditions of silencing NEDD8 or CUL1 expression (Fig. 4C), suggesting that the restoration of p27 expression was attributed to a break of CUL1-mediated ubiquitination on p27. Instead, overexpressing CUL1 significantly increased p27 ubiquitination and decreased its protein expression in sorafenib-resistant cells, which could be reversed by MLN4924(1 μM, Fig. 4C). As a result, p27, which was otherwise quickly degraded in resistant cells, was largely stabilized after MLN4924 treatment (1 μM, Fig. 4D and Supplementary Fig. 3B) or when knocking down of NEDD8 or CUL1 (Fig. 4E and Supplementary Fig. 3C), respectively.
Fig. 4
CUL1 neddylation mediates p27 ubiquitination in sorafenib-resistant liver cancer cells. (A) Western blot of SCF components (CUL1 and SKP2), NEDD8-Cullin, and p27 in sorafenib-resistant cells and their corresponding parental cells in the absence or presence of gradient neddylation inhibitor, MLN4924. (B, C) Co-immunoprecipitation analysis demonstrated the changes in the amounts of p27-immunoprecipitated CUL1 and ubiquitin under the conditions of MLN4924 treatment (1 μM) (B), NEDD8 or CUL1 silencing or overexpressing (C). (D, E) Western blot showed increased p27 protein stability in sorafenib-resistant cells under the conditions of MLN4924 treatment (1 μM) (D), NEDD8 or CUL1 silencing (E)
Hyperactive autophagy in sorafenib-resistant liver cancer cellsAs shown in the results in Fig. 1F, CUL1, SKP2 and NEDD8 were linked to autophagy receptor, NBR1. Apart from it, several genes closely related to autophagy, such as SQSTM1(also known as p62), OPTN and NDP52 were also increased in tumor tissues compared to normal tissues in GSE144269 and TCGA datasets (Fig. 5A). Moreover, NBR1, SQSTM1, OPTN and NDP52 mRNA were increased in the sorafenib-resistant tumor tissues compared to the primary tumor tissues in GSE144269 datasets. The increased NBR1 and SQSTM1 mRNA were further identified in the sorafenib-resistant tissues compared to the primary tissues (Fig. 5B). However, except the increased protein levels of OPTN, the protein levels of NBR1 and SQSTM1 were decreased in the sorafenib-resistant cells (Fig. 5E and Supplementary Fig. 4A) and tissues (Fig. 5C and D). Therefore, we conjectured that the increased NBR1 and SQSTM1 mRNA contributes to their protein levels and the formation of autophagosomes in early autophagy, but the degradation of autophagosomes in lysosomes in later autophagy leads to the final reduction of NBR1 and SQSTM1 proteins. To validate the conjecture, we used the mCherry-/eGFP-LC3B dual-fluorescent autophagy indicator system to assess autophagic state. In detail, yellow spots represent autophagosomes, and the red spots represent autolysosomes (the fusion of autophagosomes with lysosomes) (Hua et al. 2015). The microscope imaging showed that sorafenib-resistant cells had hyperactive autophagy compared to the parental cells, as evidenced by the enhanced yellow and red spots (Fig. 5F). Autophagy inhibitor Bafilomycin A1 (BafA1) blocks the formation of autophagic lysosomes and prevents the degradation of autophagy receptor proteins (Mauvezin and Neufeld 2015). The application of BafA1 induced the formation of autophagosomes but prevented their subsequent fusion with lysosomes, as indicated by the increased yellow spots and decreased red spots. Rapamycin (RAP) is an autophagy activator (Huang et al. 2015; Lin et al. 2018). Treatment with RAP increased the yellow and red spots in sorafenib-resistant cells, suggesting that it further enhances the formation of autophagosomes and autolysosomes. The microtubule-associated proteins light chain 3B (LC3B) is important in the formation of autophagosomes. The western blot results showed that LC3B-II was upregulated in both sorafenib-resistant cells than in their parental cells (Fig. 5G and Supplementary Fig. 4B). RAP treatment increased both LC3B-I and LC3B-II in sorafenib-resistant cells, which was similar to the effect by BafA1. However, RAP treatment further decreased NBR1 and SQSTM1 proteins in the sorafenib-resistant cells, while BafA1 increased the protein levels of NBR1 and SQSTM1. These results suggested that NBR1 and SQSTM1 are indeed degradated in the lysosomes in the sorafenib-resistant cells due to the enhanced autophagy.
Fig. 5
Hyperactive autophagy in sorafenib-resistant liver cancer cells. Box plots of autophagy receptors in normal tissues, primary tumor tissues, sorafenib responders and non-responders in GSE109211 and TCGA cohorts. (B) qPCR analysis of SQSTM1 and NBR1 in adjacent normal tissues, primary tumor tissues without sorafenib treatment, and primary tumor tissues resistant to sorafenib. (C) Western blot of the SQSTM1 and NBR1 in adjacent normal tissues, primary tumor tissues without sorafenib treatment, and primary tumor tissues resistant to sorafenib. (D) Representative IHC images of NBR1 in HCC patients. (E) Western blot of the autophagy receptors in HepG2-P, Huh7-P, HepG2-SR and Huh7-SR cells. (F) Representative immunofluorescence images captured in the mCherry-/eGFP-LC3B dual-fluorescent autophagy indicator system. Yellow spots represent autophagosomes, and the red spots represent autolysosomes. (G) Western blot of LC3B (autophagic activity marker), autophagy receptors and the cyclin-dependent inhibitor p27 in parental and resistant cell lines with or without BafA1 and Rapamycin treatments. Rapamycin (RAP) was used as an autophagy activator. *P < 0.05, **P < 0.01 and ***P < 0.001 (n = 3)
p27 was largely decreased in the resistant cells. After the intervention of BafA1, the protein expression of p27 was upregulated (Fig. 5G and Supplementary Fig. 4B). Instead, RAP further decreased p27 in the resistant cells. These results suggested that p27 protein level is related to the enhanced autophagy in the sorafenib-resistant cells. Notely, the qPCR data showed no between-group variance in p27 expression, while CUL1 and SKP2 mRNA were upregulated in all resistant cell groups (Supplementary Fig. 5), indicating that the expression changes of p27 did not occur at the transcription level.
Cul1 induces K29-linked ubiquitination on p27, resulting in the autophagic degradation in sorafenib-resistant liver cancer cellsAutophagy is a lysosome-degradation program where the double-membrane autophagosomes are formed in the cytoplasm to sequester cellular components (Chen et al. 2019a). Considering the hyperactive autophagic state of sorafenib-resistant cells (Figs. 3E-H), we postulated that CUL1 neddylation-mediated ubiquitination leads to the autophagy-mediated lysosomal degradation of p27 in the resistant cells, though ubiquitination usually induces the degradation of p27 in proteasomes. Previous reports have shown that MLN4924 (1 μM) induces autophagy in liver cancer cells (Luo et al. 2012). Our western blot assay showed a higher level of LC3B in MLN4924-treated sorafenib-resistant cells than untreated cells (Fig. 6A). Consistent with this, microscope imaging data also indicated an increased formation of autophagosomes in the presence of MLN4924 (1 μM, Fig. 6B). However, little or no difference was observed in terms of the amount of autophagic lysosomes, as evidenced by an increase in yellow but not red spots in MLN4924-treated cells (Fig. 6B). These results together revealed that MLN4924, a NEDD8-activating enzyme inhibitor, can promote autophagosome formation, but fail to promote autophagic flux effectively.
Fig. 6
CUL1 induces K29-linked ubiquitination on p27, resulting in the autophagic degradation in sorafenib-resistant liver cancer cells (A) Western blot of autophagic activity marker LC3B in sorafenib-resistant liver cancer cells upon MLN4924 treatment (1 μM). (B) Immunofluorescence images demonstrated increased formation of autophagosomes but not the amount of autophagic lysosomes in MLN4924-treated cells (1 μM). (C) Western blot of p27 protein in sorafenib-resistant cells in the presence of BafA1 or MG132. (D) Western blot of p27 protein in sorafenib-resistant cells when silencing the autophagy receptors. (E) Immunofluorescence images showed abrogated basal autophagy by knocking down SQSTM1 in sorafenib-resistant cells, which was reversible upon over-expressing NRB1. (F) Western blot of p27 protein in SQSTM1 knockdown cells when over-expressing NBR1 with or without BafA1 treatment. (G) HepG2-SR cells were transfected with wild and mutant types of HA-labeled ubiquitin. For the mutant ubiquitins, polyubiquitin chains are linked only at one of the lysine residues (K27, K29, K33, K48 and K63) as the other lysine residues are mutant to arginine. In addition, cells were transfected with FLAG-labeled p27 with or without CUL1 knockdown. Co-immunoprecipitation was conducted to determine the types of polyubiquitin chains on p27. (H) HepG2-SR cells were transfected with mutant types of HA-labeled ubiquitin, K27, K29, K29R and K48R. For the mutant K27 and K29 ubiquitins, polyubiquitin chains are linked only at K27 and K29 respectively, as the other lysine residues are mutant to arginine. For mutant K29R and K48R ubiquitin, the polyubiquitin chains can be linked by any lysine residues, except from the K29 and K48 respectively. In addition, cells were transfected with FLAG-labeled p27 and HIS-labeled CUL1. Co-immunoprecipitation was conducted to determine the types of polyubiquitin chains on p27. *P < 0.05, **P < 0.01 and ***P < 0.001 (n = 3)
To distinguish between autophagy- and ubiquitination-mediated p27 protein degradation, we pre-treated sorafenib-resistant cells with BafA1 or proteasome inhibitor MG132. The former blocks the formation of autophagic lysosomes (Yang et al. 2021), while the latter inhibits ubiquitin–proteasome system-mediated protein degradation (Lee and Goldberg 1998). Next, the cells were explored to the protein synthesis inhibitor cycloheximide. Finally, p27 expression at different time points were examined using western blot assay. It was discovered that p27 protein was quickly degraded in untreated cells (Fig. 6C and Supplementary Fig. 6A). However, BafA1 and MG132 were able to extend the stability of p27 protein, with BafA1 exhibiting a stronger stabilizing effect than MG132 (Fig. 6C and Supplementary Fig. 6A). Thus, the autophagy, which is activated after gaining drug-resistance, plays more important role in p27 protein degradation.
The expression of autophagy receptors (SQSTM1, NBR1, and OPTN) was different in the drug-resistant tissues and cells compared to the primary tissues and cells (Fig. 5E). Therefore, they might be implicated the autophagic degradation of p27. To validate it, we knocked down each individual receptor in the resistant cells. The results showed that p27 expression was largely induced after NBR1 knockdown, and to a less great extent upon SQSTM1 knockdown (Fig. 6D and Supplementary Fig. 6B). Silencing of OPTN showed little influence on p27 expression (Fig. 6D and Supplementary Fig. 6B). Consistently, SQSTM1 knockdown greatly abrogated high basal level of autophagy in sorafenib-resistant cells by inhibiting autophagic lysosome formation (Fig. 6E). In detail, there was increased yellow spots with decreased red spots in SQSTM1 knockdown cells versus the untreated cells (Fig. 6E). Interestingly, overexpression of NBR1 rescued the reduced autophagic flow in SQSTM1 knockdown cells, as evidenced by more prominent red spots in dual-fluorescent autophagy indicator system (Fig. 6E) and the reduced p27 protein levels in western blot assay (Fig. 6F and Supplementary Fig. 6C). Moreover, ectopic NBR1 expression alone was sufficient to induce autophagic flow according to the increased red to yellow spots ratio (Fig. 6E) and p27 protein expression (Fig. 6F and Supplementary Fig. 6C). This effect could be abrogated in the presence of BafA1, the inhibitor of autophagic lysosomes formation (Fig. 6F and Supplementary Fig. 6C).
Ubiquitin is a protein of 76 amino acids with 7 lysine residues (K6, K11, K27, K29, K33, K48, and K63), all of which can be ubiquitinated to form ubiquitin polyubiquitin chains. Different polyubiquitin chains determine the fate of substrate proteins, such as whether the protein is degraded by autophagic lysosomes or ubiquitin–proteasome system. To determine the types of polyubiquitin chains on p27, we transfected HepG2-SR cells with wild and mutant types of HA-labeled ubiquitin. For the mutant ubiquitins, polyubiquitin chains are linked only at one of the lysine residues (K27, K29, K33, K48 and K63) as the other lysine residues are mutant to arginine. As indicated by co-immunoprecipitation, the polyubiquitin chains on p27 can be linked by K27, K29, K33, K48 and K63 (Fig. 6G). However, knockdown of CUL1 decreased the polyubiquitin chains linked by K29 on p27 (Fig. 6G). We also transfected HepG2-SR cells with mutant types of HA-labeled ubiquitin, K29R and K48R (Fig. 6H). For two mutant ubiquitin, the polyubiquitin chains can be linked by any lysine residues, except from the K29 and K48 respectively. Overexpression of His-labeled CUL1 induced the addition of the K29-linked polyubiquitin chains to p27, but had minor effect once K29 was mutant to K29R. Overexpression of His-labeled CUL1 also induced the addition of the K48-linked polyubiquitin chains to p27, which is not influenced by the mutation of K48 to R48. These results collectively suggest that CUL1 primarily induces K29-linked ubiquitination on p27.
This study conducted Co-IP to determine whether K29-linked ubiquitination on p27 contributes the binding of p27 to NBR1 and SQSTM1. HA-labeled ubiquitin (K29) and Flag-label p27 vectors were transfected to HepG2-SR cells. Knockdown of CUL1 decreased the polyubiquitin chains linked by K29 on p27. Importantly, the enrichment of NBR1 and SQSTM1 binding to p27 was also decreased with CUL1 knockdown (Supplementary Fig. 6D).
Blocking neddylation and CUL1-NBR1 restores drug sensitivity in sorafenib-resistant cellsSince CUL1 neddylation and subsequent CUL1-NBR1-mediated autophagic degradation significantly influences p27 protein level, we investigated that whether restoring p27 expression by suppression of neddylation and knockdown of CUL1 affects the development of sorafenib resistance in liver cancer cells. We observed that the sensitivity of the sorafenib was regained by either adding neddylation inhibitor MLN4924 (1 μM) or knocking down CUL1 in sorafenib-resistant cells in terms of cellular proliferation (Fig. 7A and Supplementary Fig. 7A), colony-formation (Fig. 7B and Supplementary Fig. 7B) and invasion (Fig. 7C). These interventions were also able to prevent p27 degradation in the resistant cells (Fig. 7D and Supplementary Fig. 7C). Importantly, the combination of MLN4924 (1 μM) and CUL1 knockdown resulted in the strongest suppression of growth, colony-formation and invasion, as well as the highest p27 protein levels in the presence of sorafenib. However, the regained drug sensitivity of sorafenib-resistant cells achieved by CUL1 knockdown was reversible when overexpressing NBR1 and knocking down p27, as indicated by the enhanced cell growth (Fig. 7E and Supplementary Fig. 7D), colony-formation (Fig. 7F and Supplementary Fig. 7E) and invasion (Fig. 7G), and decreased p27 protein levels (Fig. 7H). CUL1 knockdown reduced the immunofluorescence of p27 merged with the immunofluorescence of lysosomes, suggesting that CUL1 knockdown suppressed the transport of p27 to lysosomes (Fig. 7I). However, overexpressing NBR1 attenuated the effect by CUL1 knockdown.
Fig. 7
Blocking neddylation and Cul1-NBR1 restores drug sensitivity in sorafenib-resistant cells (A) EdU incorporation assay showing reduced proliferation or activity of sorafenib-resistant cells under the conditions of CUL1 knockdown or MLN4924 treatment (1 μM). (B) Colony formation assay demonstrating decreased colony formation of sorafenib-resistant cells culture under the conditions of CUL1 knockdown or MLN4924 treatment (1 μM). (C) Transwell assay demonstrating inhibited invasion of sorafenib-resistant cells under the conditions of CUL1 knockdown or MLN4924 treatment (1 μM). (D) Western blot analysis demonstrating changes in p27 protein levels under conditions of CUL1 silencing with or without simultaneous addition of MLN4924 (1 μM). (E) EdU incorporation, (F) Colony formation and (G) Transwell assays show increased proliferation, colony formation and invasion when over-expressing NBR1 or knocking down p27 in CUL1 knockdown sorafenib-resistant cells. (H) Western blot showing re-stabilized p27 protein when over-expressing NBR1 in CUL1 knockdown sorafenib-resistant cells. (I) CUL1 knockdown reduced the immunofluorescence of p27 merged with the immunofluorescence of lysosomes. However, overexpressing NBR1 attenuated the effect by CUL1 knockdown. (J and K) Tumor volume and growth images demonstrating largely reduced xenografted tumor volume by day 21 after subcutaneous injection of CUL1 knocked down sorafenib-resistant HepG2 cells. (L) Representative immunohistochemistry images showing reduced proliferating marker Ki67 and increased p27 protein expression in CUL1 knocked down sorafenib-resistant HepG2 cell-derived xenografted tumors. *P < 0.05, **P < 0.01 and ***P < 0.001 (n = 3 in study in vitro and n = 6 in study in vivo)
Finally, we examined the therapeutic efficacy in vivo using the xenografted mouse model. As expected, subcutaneous injection of sorafenib-resistant HepG2 cells resulted in apparent tumor growth in the transplanted mice (Fig. 7J and K). Every other day-gavage delivery of sorafenib ameliorates the tumor burden brought by the drug-resistant cells. Knockdown of CUL1 largely sensitized cells’ response to sorafenib treatment, leading to a greatly reduced xenograft tumor size. Moreover, large tumors in the drug-resistant HepG2 transplanted mice exhibited strong nuclear immunostaining of proliferation marker proliferating cell nuclear antigen (ki67) but weak p27, regardless of sorafenib administration. In contrast, CUL1 knockdown tumors showed little ki67 staining but more strikingly increased p27 staining (Fig. 7L).
Comments (0)