
Remember me
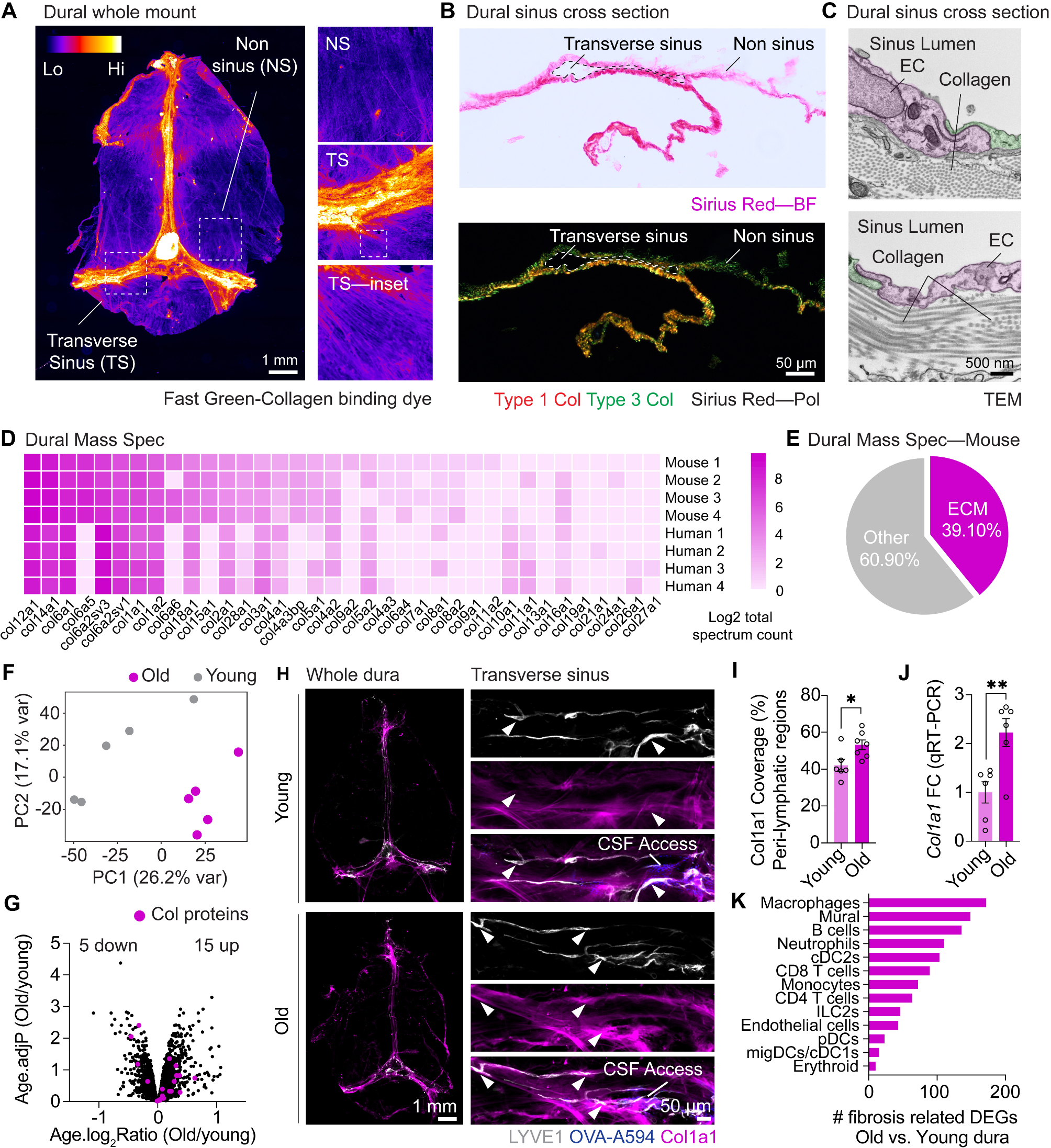


Significant decreases in the rate of weight gain (Fig. 2B) of CMU infected mice compared to SPG infected mice were observed in neonates from days 6 to 15 (endpoint), with the exception of day 12, as shown in Fig. 2B. A significant decrease in weight gain is consistent with infection in mice.
Fig. 2
Acute effects of a neonatal respiratory infection on weight. The A) average gross weight and B) weight gain per day of SPG (solid line) and CMU (dotted line) infected mice, with significance between weight gain on a single day, SPG n = 27, CMU n = 28 (*p < 0.05, **p < 0.01, ****p < 0.0001)
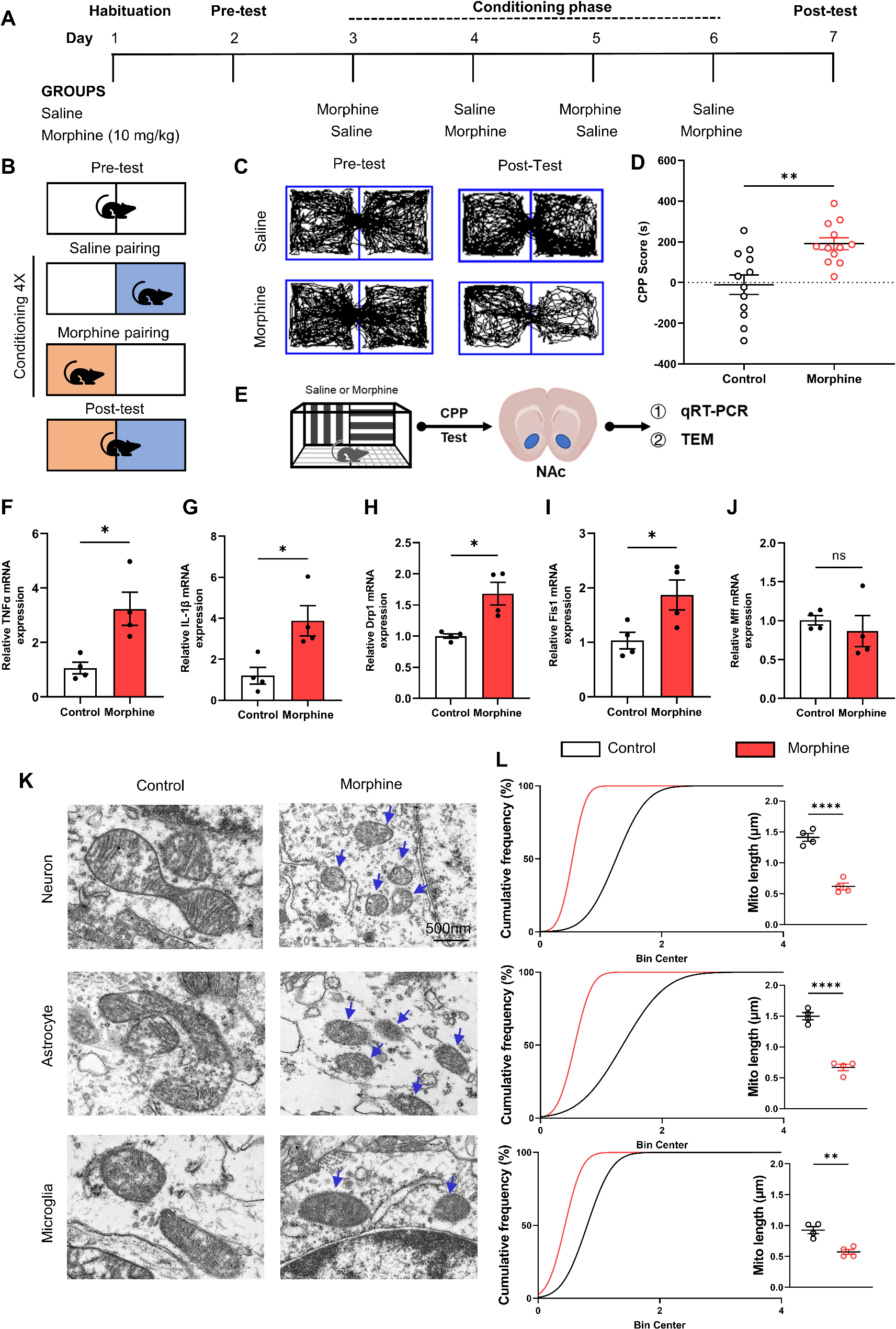
Confirmation of Chlamydia muridarum infection in the lungsAlong with the weight data to confirm infection, the expression of 16 S mRNA was measured at P15 in the lungs. The expression of a number of key inflammatory mediators were also quantified in the lungs to ensure that peripheral inflammation was present.
16 S is the central structural component of the bacterial subunit that is conserved in bacteria and allows identification of bacteria [21]. Given the variability in bacterial titre and clearance, a non-parametric test was performed, as 16 S expression data failed tests of homogeneity and Levene’s test of equal variances. There was a significant main effect of treatment, as shown in Fig. 3A, (treatment effect; H (1,21) = 21.06, p < 0.001), this indicates that both male and female cohorts were successfully infected. Chemokine ligand 9 (CXCL9) is thought to aid in the infiltration and retention of activated T-cells in response to bacterial infection [22] and will give an indication of the direct response to the bacterial infection. There was a significant main effect in the expression of CXCL9 in the lungs at P15, shown in Fig. 3B (treatment effect; H (1,25) = 21.85, p < 0.001). This indicates that the initial peripheral response to the bacterial infection is not sex-specific.
NLR family pyrin domain containing 3 (NLRP3) is a major component of the inflammasome and triggers the immune response. Once triggered, NLRP3 activates caspase-1 which then cleaves pro-interleukin 1β into its mature form, the a highly pro-inflammatory interleukin 1β (IL-1β) [23]. Tumour necrosis factor alpha (TNFα) is a pro-inflammatory cytokine produced during acute inflammation [24]. These mediators act together to initiate the inflammatory response in the lungs. At P15, there was a main effect of treatment on NLRP3, TNFα and IL-1β (treatment effect, NLRP3; F (1,25) = 45.229, p < 0.001, TNFα; F (1,25) = 13.235, p = 0.001, IL-1β; F (1,25) = 51.409, p < 0.001) with CMU-infected mice having a significant increase compared to SPG-infected mice, as shown in Fig. 3C, D, E. There was no other main effects or interactions. This indicates that in the lungs there is a robust inflammatory response to infection that is not sex-specific.
Fig. 3
Upregulation in the expression of all inflammatory mediators in the lungs following neonatal infection. Graphs show gene expression levels (mean ± SEM) for SPG (solid bars) and CMU (hatched bars) females (grey) and males (black), with total number of samples shown in each bar. Inflammatory mediator expression; A) 16s (SPG vs. CMU ***p < 0.001), B) CXCL9 (SPG vs. CMU ***p < 0.001), C) NLRP3 (SPG vs. CMU ***p < 0.001), D) TNFα (SPG vs. CMU **p < 0.01) and E) IL-1β (SPG vs. CMU ***p < 0.001)
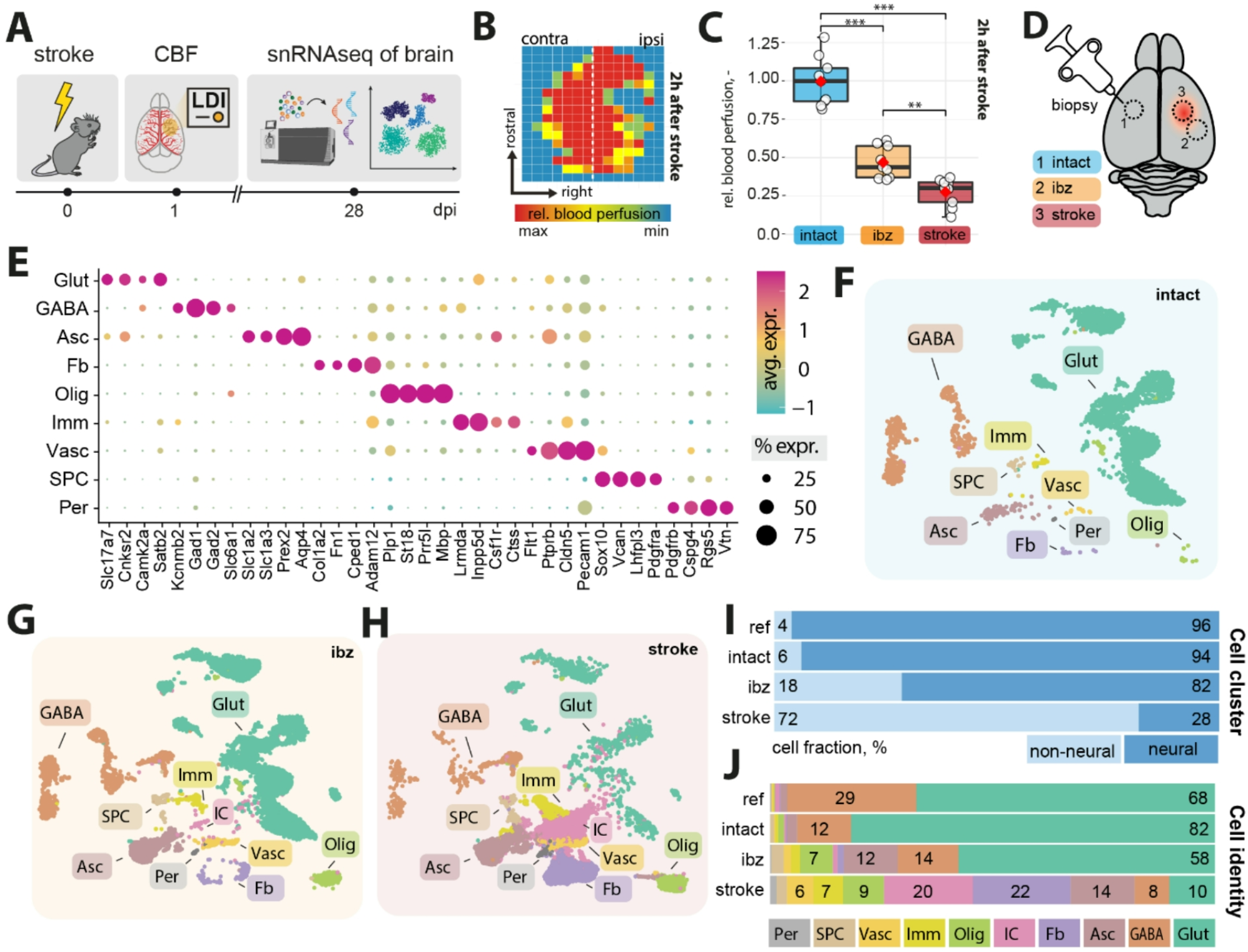
Neonatal infection alters inflammatory mediators in the medulla oblongataThis study examined the expression of a wide range of inflammatory mediators in the medulla oblongata in the acute phase of peripheral inflammation. Inflammatory mediators selected have been shown to be upregulated peripherally in response to infection, as well as mediators that a play crucial role in regulating the local immune environment of the CNS.
Upregulation of neuroprotective inflammatory mediators in infected femalesCXCL9 and chemokine ligand 10 (CXCL10) are two major chemokines that have been associated with immune cell migration, differentiation and activation [25]. Specifically, CXCL10 is primarily expressed on glia and recruits lymphocytes and phagocytes to inflamed areas to limit the spread of infection [26, 27] and as is the case with CXCL9, aids in T-cells responses to bacterial infection. These two chemokines will indicate the strength of the initial immune response to the bacteria. At P15 there was a significant treatment x sex effect in the expression of both CXCL9 and CXCL10 (treatment x sex, CXCL9; F (1,26) = 6.682, p = 0.016, CXCL10; F (1,27) = 5.472, p = 0.027). FCMU had a significant increase in the expression of both CXCL9 and CXCL10 compared to FSPG, as shown in Fig. 4A, B (post-hoc for females, CXCL9; F (1,26) = 15.251, p = 0.001, CXCL10; F (1,27) = 4.843, p = 0.036). There was no significant difference in MCMU compared to MSPG (post-hoc for males, CXCL9; F (1,26) = 0.062, p = 0.805, CXCL10; F (1,27) = 0.024, p = 0.879). These results suggest that females, unlike their male counterparts, are mounting a stronger response within the CNS to inhibit the development of infection during the acute phase.
Interleukin-13 (IL-13), functions in the CNS as an anti-inflammatory cytokine and is expressed by reactive microglia and can produce both beneficial and harmful effects on the local environment [28]. The IL-13 inflammatory pathway has been shown to modulate potassium calcium-activated channel subfamily N member 4 (KCNN4) which codes for KCa3.1 [29]. KCa3.1 is a voltage-independent potassium channel that is activated by intracellular calcium. Activation of this channel helps to sustain calcium entry into cells via voltage independent pathways and is critical for the inflammatory response in microglia and astrocytes [30, 31].
At P15, treatment x sex analysis revealed sex-specific effects of infection on both of these inflammatory mediators (treatment x sex, IL-13; F (1, 26) = 5.820, p = 0.026, KCNN4; F (1, 27) = 7.661, p = 0.010). For the expression of IL-13, there was significant difference between FCMU and FSPG, as shown in Fig. 4C (post-hoc for females; F (1,26) = 5.281, p = 0.030) while there were no differences between males (post-hoc for males; F (1,26) = 0.245, p = 0.624). Similarly, KCNN4 expression also showed significant differences between FCMU and FSPG, as shown in Fig. 4D (post-hoc for females; F (1,27) = 6.192, p = 0.019) while there were no differences between males (post-hoc for males; F (1,27) = 1.982, p = 0.171). This indicates that there are significant changes within the IL-13 inflammatory pathway in females only during the acute inflammatory response to a peripheral respiratory infection.
Brain derived neurotrophic factor (BDNF) is a crucial component in brain plasticity, neuronal development, growth and survival, and is implicated in establishing functional neuronal networks during development [32] .Transforming growth factor beta (TGFβ) is a multifactorial cytokine that regulates the behaviour of many immune cells, such as macrophages and neutrophils, causing anti-inflammatory effects and forming a network of negative immune regulatory inputs [33]. TGFβ signalling can regulate BDNF and both are considered protective when acutely responding to inflammation [34]. At P15, treatment x sex analysis revealed a significant effect of infection on BDNF expression (Fig. 4E), but no change to the expression of TGFβ (Fig. 4F) (treatment x sex, BDNF; F (1, 27) = 5.235, p = 0.031, TGFβ; F (1, 27) = 4.009, p = 0.055). The expression of BDNF in FCMU was significantly increased compared to FSPG, shown in Fig. 4E (post-hoc for females; F (1,27) = 5.054, p = 0.033) and there was no significant difference in males (post-hoc for males; F (1,27) = 0.056, p = 0.815). Together, this suggests that the initial potential protective response in the medulla oblongata is only seen in females, and it is driven by BDNF expression, not TGFβ.
Fig. 4
Upregulation of inflammatory mediators in the medulla oblongata of infected females. Graphs show A) CXCL9 (FSPG vs. FCMU **p < 0.01), B) CXCL10 (FSPG vs. FCMU *p < 0.05), C) IL-13 (FSPG vs. FCMU *p < 0.05), D) KCNN4 (FSPG vs. FCMU *p < 0.05), E) BDNF (FSPG vs. FCMU *p < 0.05) and F) TGFβ relative gene expression levels (mean ± SEM) at P15. Graphs presented as SPG (solid bars), CMU (hatched bars), females (grey) and males (black), with total number of samples shown in each bar
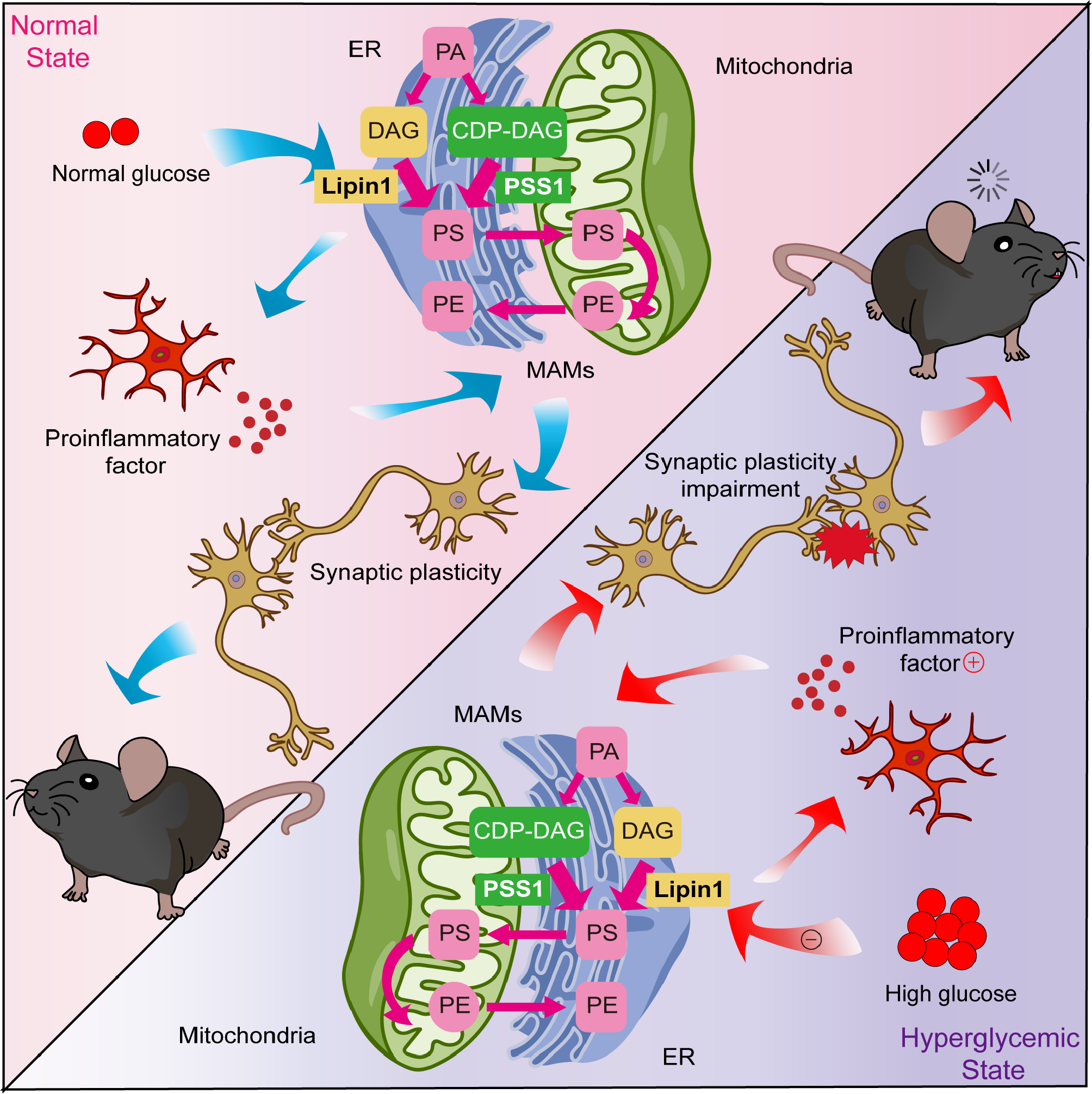
Suppression of inflammatory mediators in infected malesSignal transducer and activator of transcription 6 (Stat6) is required for the promotion of the T-helper type 2 (Th2) immune response [35]. There was a significant treatment x sex interaction for Stat6 (treatment x sex, Stat6; F (1, 27) = 5.831, p = 0.023). The expression of Stat6 showed a significant decrease in MCMU compared to MSPG, as shown in Fig. 5A (post-hoc for males; F (1,27) = 5.395, p = 0.028) while there were no differences between females (post-hoc for females; F (1,27) = 0.017, p = 0.896). This indicates that there are significant sex-specific changes, with infected males showing a suppression of potential anti-inflammatory pathways in the acute inflammatory response to a peripheral respiratory infection.
Prostaglandin-endoperoxide synthase 2 (PTGS2), also known as cyclooxygenase (COX-2), is a key enzyme in prostaglandin biosynthesis and during inflammation is associated with oedema and leukocyte trafficking [36]. Treatment x sex analyses revealed a significant effect on the expression of PTGS2 (Fig. 5B, treatment x sex, PTGS2; F (1, 26) = 5.439. p = 0.028). MCMU showed a significant decrease compared to MSPG (post-hoc for males; F (1,26) = 4.347, p = 0.047), with no differences seen in females (post-hoc for females; F (1,26) = 1.472, p = 0.236). This indicates a clear suppression of male PTGS2 in response to the infection.
Interferon gamma (IFNγ) is a pro-inflammatory cytokine that primarily activates macrophages and stimulates neutrophils [37]. Signal transducer and activator of transcription 1 (Stat1) increases macrophage inflammatory responses [38] and is one of the primary transcription factors activated by IFNγ, thus playing a major role in immune responses [39]. However, at P15 no significant differences were seen in either IFNγ or Stat1, shown in Fig. 5C, D, indicating that, at this timepoint, this pathway is not heavily involved in the medulla oblongata’s initial response to a neonatal bacterial infection.
Given the importance of NLRP3, IL-1β, TNFα in the acute phase of the peripheral inflammatory response, the expression of these inflammatory mediators was also assessed in the medulla. However, there were no significant differences in NLRP3, TNFα or IL-1β, shown in Fig. 5E-G, indicating that, at this acute timepoint, this pathway is not heavily involved in the medulla oblongata’s acute response to a neonatal infection.
Fig. 5
Downregulation of inflammatory mediators in the medulla oblongata of infected males. Graphs show A) Stat6 (MSPG vs. MCMU *p < 0.05), B) PTGS2 (MSPG vs. MCMU *p < 0.05), C) IFNγ, D) Stat1, E) NLRP3, F) TNFα and G) IL-1β relative gene expression levels (mean ± SEM) at P15. Graphs presented as SPG (solid bars), CMU (hatched bars), females (grey) and males (black), with total number of samples shown in each bar
Glia in the respiratory centres of the brainstem are significantly affected by a neonatal respiratory infectionBoth microglia, labelled with Iba1, and astrocytes, labelled with GFAP, were assessed in key regions of the medulla oblongata known to be important to the central processing of respiratory function. These are the nucleus tractus solitarii (NTS) and dorsal motor nucleus of the vagus (DMX).
NTS and DMX show significant alterations to microglia activation and morphologyIn the NTS, there was a main effect of treatment on the number of cells (Fig. 6E; treatment effect; F (1,18) = 23.946, p < 0.001). However, when this was further explored through treatment x sex analysis, a significant difference was only present between the number of cells in FCMU and FSPG (post-hoc for females; F (1,18) = 23.815, p < 0.001), whereas males were approaching significance (post-hoc for males; F (1,18) = 4.163, p = 0.056). This indicates that the magnitude of change to the number of cells differs between females and males. This sex-difference was mirrored in other analyses, with treatment x sex analysis revealing a significant difference in the mean fluorescent intensity (MFI) in males, where MCMU increased compared to MSPG, shown in Fig. 6F (treatment effect; F (1,20) = 5.531, p = 0.029) (post-hoc for males; F (1,20) = 17.410, p < 0.001). In contrast, females showed a significant difference in % area, with FCMU increasing compared to FSPG, shown in Fig. 6G (treatment effect; F (1,20) = 15.109, p = 0.001) (post-hoc for females; F (1,20) = 16.089, p < 0.001). Interestingly, treatment x sex analysis of microglia in the NTS revealed differences between the sexes in % area, with FSPG being smaller than MSPG, shown in Fig. 6G (post-hoc for SPG; F (1,20) = 7.381, p = 0.013). Further morphological analyses were undertaken to examine the specific characteristics of the microglia. There was a main effect of treatment of soma area, Fig. 6H, (treatment effect; F (1,19) = 29.554, p < 0.001). Again, when treatment x sex analysis was performed a magnitude difference was observed, with a greater difference observed between MCMU and MSPG (post-hoc for males; F (1,18) = 21.919, p < 0.001, post-hoc for females; F (1,18) = 9.039, p = 0.007). There was also a treatment x sex effect on the number of secondary branches (branch points), with FCMU decreasing compared to FSPG (treatment effect; F (1,18) = 3.612, p = 0.073) (post-hoc for females; F (1,18) = 5.076, p = 0.037), with no difference in males. Taken all together, the female’s response to infection results in more microglia, with larger somas and retracted branches, which accounts for the increase in % area. Whereas the male’s response to infection results in a bigger soma and a greater production of Iba1, as shown by the MFI. This indicates that while both female and male microglia are being affected in the acute phase of infection, the characteristics of the response to the insult differs.
Fig. 6
Iba1 immunofluorescent images and quantification of microglia in the NTS. Brainstem sections were labelled with Iba1 to identify microglia and nuclei were identified using a brain atlas. A-D) representative images of all groups showing the area selected for NTS with the area postrema removed, scale bars represent 100 μm. Graphs show E) number of cells (FSPG vs. MCMU ***p < 0.001), F) MFI (MSPG vs. MCMU ***p < 0.001), G) % area (FSPG vs. FCMU ***p < 0.001, FSPG vs. MSPG *p < 0.05), H-J) units in pixels, with H) soma area (FSPG vs. FCMU **p < 0.01, MSPG vs. MCMU ***p < 0.001), I) number of primary branches and J) number of secondary branches (branch points) (FSPG vs. FCMU *p < 0.05), (mean ± SEM). Graphs presented as SPG (solid bars), CMU (hatched bars), females (grey) and males (black), with total number of samples shown in each bar
In the DMX, treatment x sex analysis (treatment x sex, F (1, 18) = 17.458, p = 0.001) revealed there was a significant decrease in the cell number of FCMU compared to FSPG, shown in Fig. 7E (post-hoc for females; F (1,18) = 5.517, p = 0.030) while there was a significant increase in cell number of MCMU compared to MSPG (post-hoc for males; F (1,18) = 12.674, p = 0.002). This was reflected in the analysis of MFI (treatment x sex, F (1, 20) = 30.167.
p < 0.001), with a significant increase in the MFI between MCMU and MSPG, shown in Fig. 7F (post-hoc for males; F (1,20) = 22.062, p < 0.001), whereas in females, FCMU saw a significant decrease compared to FSPG, Fig. 7F (post-hoc for females; F (1,20) = 9.428, p = 0.006). Unlike the NTS, morphological analyses of the DMX showed no significant effect of infection on any characteristic, indicating in the acute phase a respiratory infection only impacts the number of microglia present in the DMX. Interestingly, sex x treatment analysis revealed differences between the sexes in soma area, with FSPG having a smaller soma than MSPG, shown in Fig. 6H (sex effect; F (1,18) = 9.810, p = 0.006) (post-hoc for SPG; F (1,18) = 4.513, p = 0.048). This indicates a different starting point between male and female microglia regardless of environmental factors.
Fig. 7
Iba1 immunofluorescent images and quantification of microglia in the DMX. Brainstem sections were labelled with Iba1 to identify microglia and nuclei were identified using a brain atlas. A-D) representative images of all groups showing the area selected for DMX, scale bars represent 100 μm. Graphs show E) number of cells (FSPG vs. FCMU *p < 0.05, MSPG vs. MCMU **p < 0.01), F) MFI (FSPG vs. FCMU **p < 0.01, MSPG vs. MCMU ***p < 0.001), G) % area, H-J) units in pixels, with H) soma area (FSPG vs. MSPG *p < 0.05), I) number of primary branches and J) number of secondary branches (branch points) (mean ± SEM). Graphs presented as SPG (solid bars), CMU (hatched bars), females (grey) and males (black), with total number of samples shown in each bar
Astrocyte changes are minimal following neonatal respiratory infectionThere was a main effect of treatment (treatment effect, F (1, 19) = 27.138, p < 0.001) on the % area of NTS GFAP + ve cells, shown in Fig. 8F. This indicates that astrocytes from both sexes show similar responses to infection. There were no changes in either MFI of the NTS or either parameter within the DMX.
Fig. 8
GFAP immunofluorescent images and quantification of astrocytes in the NTS and DMX. Brainstem sections were labelled with GFAP to identify astrocytes and nuclei were identified using a brain atlas. A-D) representative images of all groups showing the area selected for NTS with the area postrema removed, G-J) representative images of all groups showing the area selected for DMX, scale bars represent 100 μm. Graphs show E) MFI and F) % area (SPG vs. CMU ***p < 0.001) in the NTS and K) MFI and L) % area in the DMX (mean ± SEM). Graphs presented as SPG (solid bars), CMU (hatched bars), females (grey) and males (black), with total number of samples shown in each bar
Comments (0)