
Remember me





Exosomes are phospholipid vesicles ranging from 30 to 150 nanometers in length that are secreted by cells to facilitate intercellular communication through biomolecular transfer [1]. Exosomes were first observed in vitro in 1983 from cultured sheep reticulocytes and were subsequently formally named ‘exosomes’ by Johnstone in 1987. Exosomes were initially considered cellular waste products but are now recognized as pivotal in signaling and ubiquitously found in bodily fluids. Exosome generation, secretion, and molecular composition intricately depend on factors such as cell type, growth status, and receptor stimulation. For instance, kinetic and compositional differences exist between exosomes from white blood cells and red blood cells. Conditions such as oxidative stress or inflammation notably alter leukocyte exosome content [2]. Bioactive compounds, such as proteins and RNAs, encapsulated in exosomes released from cells undergoing death differ from those released by normal viable cells [3].
Cell death, including regulated processes such as apoptosis, necroptosis, autophagy, pyroptosis, and unregulated necrosis triggered by external factors, is crucial for cellular, tissue, and organismal homeostasis [4]. Despite varying initiation mechanisms, each programmed cell death form significantly influences exosome release, bioactive molecule encapsulation, and intercellular communication [3]. Exosomes from different cell death pathways play distinct roles; for example, those from apoptotic cells may activate immune responses [5], while those from necrotic or pyroptotic cells can induce immune cell death and amplify inflammation [6].
This review aims to consolidate the current understanding of the mechanisms, compositions, and functions of exosomes released during programmed cell death. Investigating these aspects, particularly in the context of regulated cell death, opens new avenues for disease diagnosis and targeted therapeutics. Overall, this review emphasizes the critical role of exosomes as regulators of communication between dying and healthy cells, highlighting their importance in cellular biology.
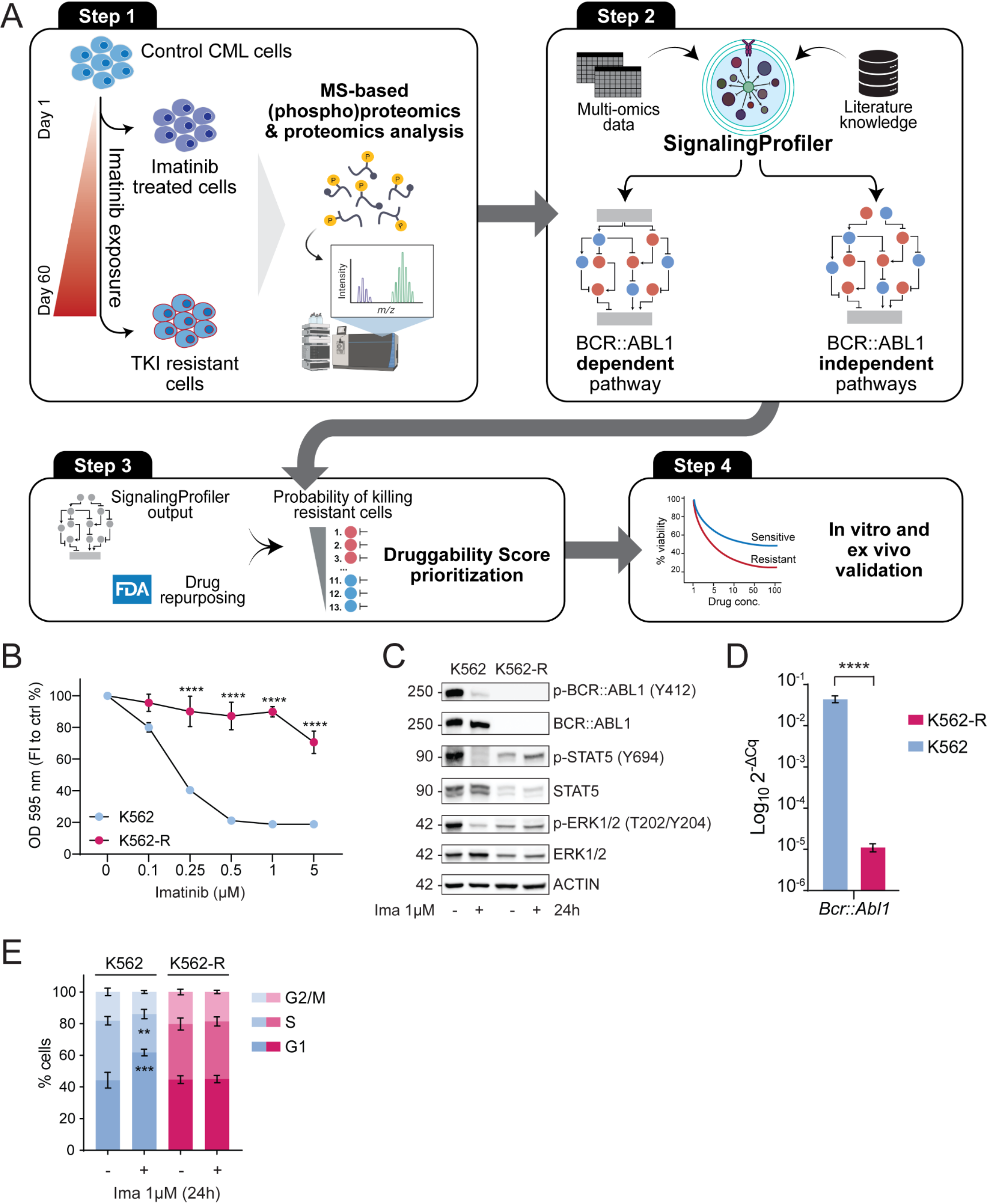
Biological functions of exosomesIntercellular communication and signal transductionExosomes, which serve as vital intercellular messengers, facilitate the conveyance of bioactive molecules to nearby target cells via paracrine secretion or distally through the circulatory system. They convey contents to recipient cells through mechanisms such as membrane fusion, receptor-mediated endocytosis, or phagocytosis. Owing to their size limitations, noncoding RNAs, including miRNAs, circRNAs, and lncRNAs, predominate within exosomes (Fig. 1). These exosomes play a significant role in regulating gene transcription in recipient cells by transporting noncoding RNAs. For instance, Jumo et al. reported that exosomes from nicotine-stimulated macrophages containing miR-21-3p promote the proliferation and migration of vascular smooth muscle cells (VSMCs) via the PTEN pathway, thereby accelerating atherosclerosis [7]. Similarly, Yin Hu et al. demonstrated that exosomes containing miR-21-3p enhance fibroblast activation and angiogenesis in skin wound healing [8]. Additionally, a considerable number of circRNAs are present in exosomes; for example, tumor-derived exosomes containing circPACRGL can augment colorectal cancer cell growth, migration, and invasion through the miR-142-3p/miR-506-3p-TGF-β axis [9].
In summary, exosomes, as minuscule cell-derived vesicles, play a crucial role in transmitting intracellular information to adjacent environments or other cells. The primary roles of these proteins in cell biology and physiology include intercellular communication and signal transduction.
Fig. 1
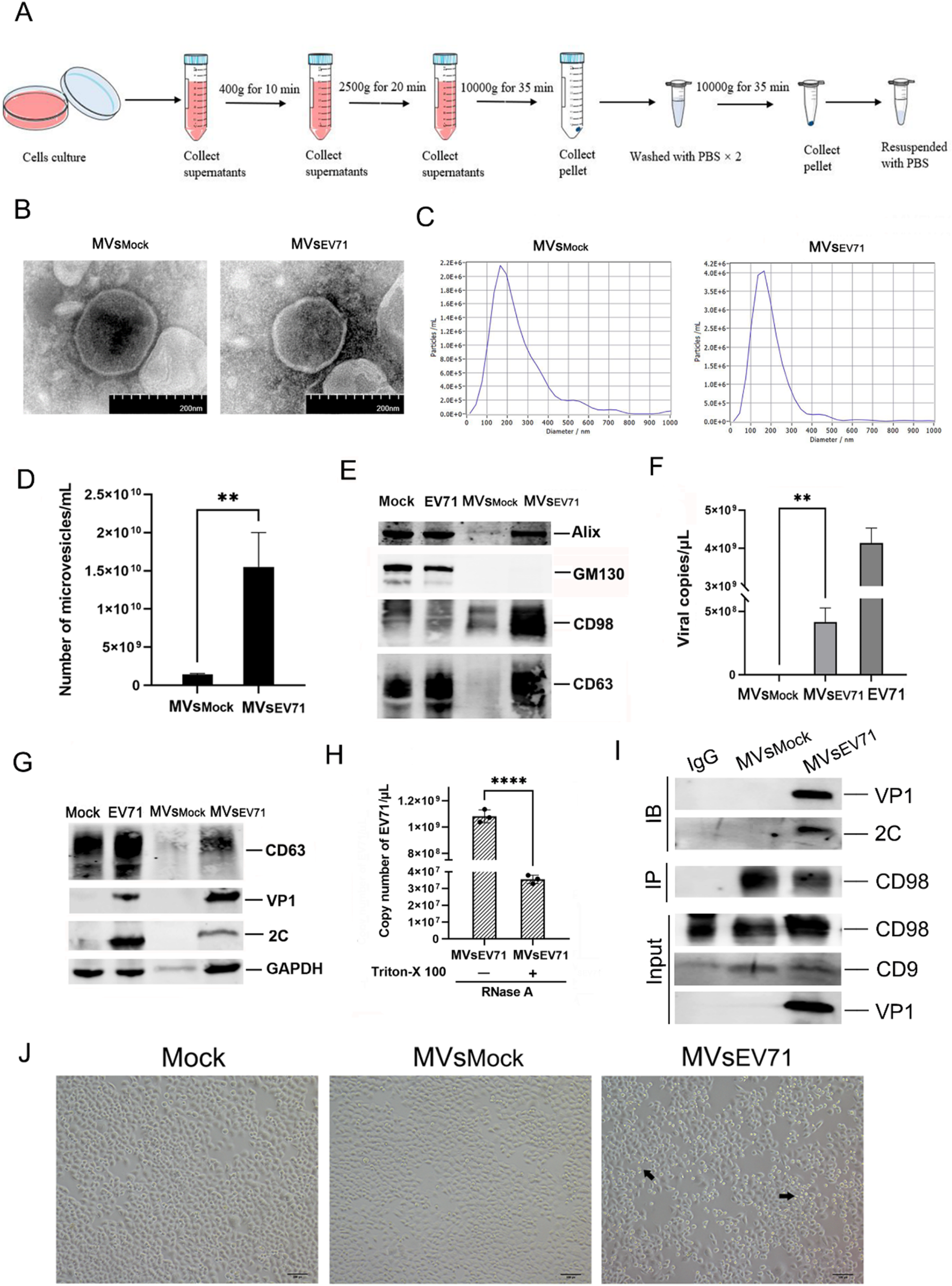
Exosome biogenesis, secretion, uptake, and isolation. A Exosome biogenesis occurs within the endosomal system and involves crucial organelles such as mitochondria, the endoplasmic reticulum, and the Golgi apparatus. It commences with plasma membrane invagination, forming early endosomes (ESEs), which subsequently fuse into late endosomes (LSEs). At this juncture, LSEs participate in cargo sorting near the Golgi apparatus, resulting in the generation of multivesicular bodies (MVBs). These MVBs can undergo lysosomal degradation, intracellular recycling, or secretion as exosomes. Once secreted, recipient cells internalize exosomes through mechanisms such as receptor-mediated endocytosis, phagocytosis, or direct fusion with the plasma membrane. B Differential centrifugation is a standard technique for isolating exosomes from conditioned cell culture media. Subsequent purification of the exosomes was achieved via ultracentrifugation at 120,000 × g. The resulting purified exosome suspension was subsequently applied to copper grids for sedimentation, followed by uranyl acetate double staining. The exosomes were subsequently visualized through transmission electron microscopy. The purified exosomes displayed a cup-shaped morphology, ranging in size from 30 to 150 nm, as observed in microscopy images (transmission electron microscopy images derived from exosomes secreted by J774A.1 macrophages extracted by our research group)
Immune regulationExosomes serve as versatile biological regulators of immune responses by employing diverse mechanisms. First, most immune cells (e.g., dendritic cells, B lymphocytes, T lymphocytes, neutrophils, and mast cells) serve as donor cells in immunoregulation by releasing exosomes carrying signaling molecules. For instance, dendritic and B-cell-derived exosomes function as antigen-presenting platforms, priming and activating T cells [10]. Notably, exosomes exhibit functional heterogeneity; those secreted by identical immune cells may exert varied impacts on the immune microenvironment, influenced by compositional differences induced by distinct stimuli. For instance, Yili Wang et al. reported that exosomes isolated from M2 macrophages, which contain mRNAs related to the PI3K/AKT pathway, could reprogram M1 macrophages to the M2 phenotype, functioning as immunomodulators and expediting diabetic fracture healing [11]. Conversely, Fuquan Zhang et al. reported that M2 macrophage-derived exosomes, which are rich in the long noncoding RNA AGAP2-AS1, enhance the efficacy of radiotherapy by inhibiting miR-296 and augmenting NOTCH2 signaling [12]. Hence, the impact of exosomes on immune regulation relies primarily on their bioactive substance content.
Moreover, exosomes facilitate downstream communication among immune cells and modulate the immune microenvironment. Specific exosomes encapsulate immunomodulatory substances such as major histocompatibility complex (MHC), costimulatory molecules, and other immunomodulators that interact with mammalian immune cell receptors, influencing their activities and functionalities [13]. Additionally, exosomes contain regulatory RNAs, including miRNAs and lncRNAs, which bind to target genes and mediate signaling pathways affecting immune cell operation and overall immune system dynamics [10]. For instance, adipose stem cell-derived exosomes can alleviate LPS-induced inflammation and provide protection against sepsis by modulating Nrf2/HO-1 expression [14]. Similarly, mesenchymal stem cell-derived exosomes can mitigate oxidative skin damage by adaptively regulating the NRF2 defense system [15]. Exosomes can also modulate innate immune responses and be engineered into therapeutic agents and vaccines for cancer, suggesting promising applications in the diagnosis, treatment, and prognosis of cancer. For instance, exosomes enriched in tumor antigens such as MHC I may serve as acellular vaccines to prime antitumor immune responses for cancer immunotherapy [16]. Moreover, exosomes harboring heat shock proteins (HSPs) and programmed death ligand 1 (PD-L1) can inhibit immune cell function and deplete the immune repertoire to facilitate immune evasion. Blocking the generation of such immunosuppressive exosomes has also emerged as a novel immunotherapeutic strategy [17].
In summary, exosomes are pivotal for immune system regulation through processes such as encapsulation, secretion, and uptake, offering novel insights and therapeutic avenues for treating immune-related disorders.
Inflammation regulationExosomes play a pivotal role in orchestrating inflammatory responses by transmitting inflammatory factors and activating downstream target cells through antigen presentation. For instance, immune cells release exosomes, which contain anti-inflammatory agents such as interleukin-10 (IL-10) and transforming growth factor-β (TGF-β), during inflammatory reactions, effectively curbing excessive inflammation [18]. Additionally, microRNAs (miRNAs) within exosomes downregulate the expression of inflammatory genes, including NF-κB and TNF-α, contributing to inflammation. Notably, bone marrow-derived macrophage exosomes carrying miR-99a/146b/378a target these pathways, reducing inflammation and inducing an anti-inflammatory M2 phenotype in recipient macrophages, thereby modulating both the intensity and duration of inflammatory responses [19].
Exosomes also exhibit dual functions in regulating inflammation. Inflammasome activation influences exosome release; conversely, exosomes can modulate inflammasome activity to either promote or suppress inflammation [20, 21]. For instance, upon exposure to zinc oxide nanoparticles (ZnONPs) and UVB radiation, pyroptotic cells release exosomes containing the NLRP3 inflammasome, extending damage to adjacent cells [22]. Overexpression of thioredoxin-interacting protein (TXNIP) activates TLR4/NF-κB/NLRP3 signaling, inducing pyroptosis in rat cardiomyocytes. However, exosomes from M2 macrophages, which carry microRNA-148a, downregulate TXNIP and impede NLRP3 inflammasome activation, thereby alleviating myocardial ischemia/reperfusion injury [23].
In summary, exosomes play a crucial role in orchestrating inflammatory responses by transporting inflammation-related factors, triggering downstream inflammatory signaling, and influencing inflammasome activity. The inflammatory impact of exosomes varies with the mode of cell death they originate from; exosomes from dying cells often exhibit more pronounced proinflammatory properties than those from viable cells. This underscores the importance of gaining a deeper understanding of how different cell death modalities and exosome release interplay in inflammation regulation, offering potential avenues for developing therapeutic strategies.
Tumor microenvironment (TME) mediationCancer cell-derived exosomes play a crucial role in mediating communication with neighboring cells, establishing a feedback loop that modulates the tumor microenvironment (TME) and fosters tumor growth, invasion, and metastasis [24]. Research by Selma Maacha et al. demonstrated that exosomes can act as carriers of carcinogens, and inhibiting their release effectively reduces carcinogen transfer [25]. Specifically, exosomes from tumors enriched with epithelial–mesenchymal transition (EMT)-related biomolecules such as TGF-β, hypoxia-inducible factor (HIF) 1α, β-catenin, and miR-23a induce EMT in cancer cells, leading to stromal remodeling and degradation [26]. This process is pivotal for tumor cell invasion, migration, and distant metastasis. Additionally, exosomes therapeutically influence the TME by impacting pathways related to intercellular communication, the immune response, matrix remodeling, and drug resistance. For example, Zhang Wei et al. reported that tumor cell-derived exosomes containing programmed cell death ligand 1 (PD-L1) can bind to programmed cell death protein 1 (PD-1) on CD8 + T cells, inhibiting T-cell activity, facilitating immune escape, and promoting tumor immune tolerance [27]. Exosomes derived from tumor cells contain various microRNAs, long noncoding RNAs, and circular RNAs that can promote tumor progression by inducing angiogenesis. However, some noncoding RNAs may conversely inhibit tumorigenesis. Targeting proangiogenic noncoding RNAs in exosomes could be a promising therapeutic strategy for cancer treatment [28]. Mesenchymal stem cell-derived exosomes reduce cancer cell drug resistance and enhance treatment efficacy [29]. Exosomes derived from mesenchymal stem cells (MSCs) can serve as excellent drug delivery vehicles for targeted cancer therapy. For instance, drugs can be loaded into purified MSC-derived exosomes (MSC-EVs) via direct insertion or coincubation methods. Engineered MSC-EVs can increase the affinity and uptake of target cancer cells. Ultimately, these optimized MSC-EVs may act as ideal drug carriers for the treatment of cancers and other diseases [30].
The development and application of therapeutic exosomes in precision medicine present promising prospects for tumor therapy, introducing innovative strategies for cancer treatment. Recent studies suggest that encapsulating exosomes with antitumor drugs or genes is an innovative approach for targeted cancer therapy [31]. Modifying the bioactive components of exosomes through genetic or pharmacological methods can selectively promote or inhibit tumor growth for therapeutic purposes. For example, exosomes loaded with miR-126 can initiate protective autophagy and programmed cell death in cancer cells, producing anticancer effects [32]. The role of exosomes in tumor progression varies with different programmed cell death pathways; for example, exosomes rich in apoptosis-inducing factor (AIF) may facilitate tumor cell apoptosis [32, 33], whereas caspase-mediated exosomes could enhance cancer cell proliferation and invasion [34, 35].
Therefore, understanding the mechanisms of exosome release in specific programmed cell death pathways and their biological functions in cancer is crucial for advancing our understanding of programmed cell death and progression in cancer diagnosis and targeted therapy.
Classification and mechanisms of programmed cell death modesApoptosisApoptosis, the earliest recognized form of programmed cell death, plays a crucial role in maintaining cellular homeostasis. It encompasses distinctive morphological alterations, including membrane reshaping, protuberances, cell shrinkage, and nuclear chromatin condensation. Apoptotic processes involve DNA fragmentation, mitochondrial dysfunction with increased permeability, and the release of cytochrome c (Cyt c) into the cytoplasm, culminating in the formation of apoptotic bodies [36]. These apoptotic bodies, which contain intracellular proteins and nucleic acids, signal neighboring cells for elimination. Phosphatidylserine exposure on apoptotic bodies allows phagocytes, such as macrophages, to engulf them. The removal of apoptotic bodies by phagocytes prevents excessive inflammation and mediates immune responses. Additionally, apoptotic cells release exosomes, a specific subclass of extracellular vesicles with distinct biogenic origins and functions compared to apoptotic bodies.
The initiation of apoptosis entails caspase enzyme activation and cascade mediation. Procaspase enzymes undergo proteolytic cleavage, activating initiator caspases (caspase-2, -8, -9, and − 10) and executioner caspases (caspase-3, -6, and − 7), leading to irreversible cell disassembly and death [37, 38]. Apoptosis can be triggered through three primary pathways: extrinsic (death ligand‒receptor binding), intrinsic (mitochondria-dependent), and endoplasmic reticulum (ER) stress-related [39].
In the extrinsic pathway, which is initiated by death ligands (FasL, TNF-α, TRAIL) binding to death receptors (FASRs, TNFR1), signaling cascades are activated, ultimately leading to apoptosis [40]. Death receptors, which have extracellular cysteine domains and intracellular death domains, form the death-inducing signaling complex (DISC), which activates caspases-8 and − 10. These caspases then activate caspase-3 and − 7, resulting in the degradation of essential cellular proteins and the induction of apoptosis [41]. Exosomes can also induce apoptosis in recipient cells through the extrinsic pathway by transporting death receptor-related molecules or regulators [42, 43].
The intrinsic mitochondrial pathway is regulated by the balance between proapoptotic and antiapoptotic proteins [44]. Apoptotic stimuli suppress antiapoptotic proteins belonging to the BCL2 family (e.g., BCL-2, BCL-xL, and MCL-1) while activating proapoptotic BH3 proteins (e.g., BID, BIM, and PUMA). This process induces conformational changes and the assembly of the BAX and BAK proteins. These proteins assemble into pore structures in the outer mitochondrial membrane, causing alterations in the mitochondrial membrane potential, increased permeability, and induction of mitochondrial outer membrane permeabilization (MOMP) [38]. MOMP leads to the release of proapoptotic mitochondrial contents, such as cytochrome c (Cyt c), into the cytoplasm, thereby facilitating apoptotic processes. Cyt c binds to Apaf-1, resulting in the formation of the apoptosome and the activation of caspase-9. Subsequently, caspase-9 triggers the executioner caspases (caspase-3, -6, and − 7) in the apoptotic cascade [44]. Activated caspase-3 not only governs apoptosis but also facilitates the release of extracellular vesicle-like exosomes (ApoExos), influencing recipient cells [45]. These exosomes may influence mitochondrial membrane proteins via miRNA release, affecting intrinsic pathway activity and mitochondrial membrane permeability, or they may directly transmit molecules such as Cyt c to modulate apoptosis [46, 47].
The endoplasmic reticulum (ER) serves as the primary intracellular organelle for calcium storage, protein synthesis, and protein modification. Elevated ER stress, resulting from calcium imbalance or the accumulation of misfolded proteins, triggers apoptosis via the ER stress-induced cell death pathway [48]. Furthermore, ER stress facilitates the release of exosomes, which subsequently modulate the activity of the ER stress pathway by influencing molecules such as PERK and ATF6 [49, 50].
NecroptosisNecroptosis, triggered by severe stimuli such as hypoxia, viral infection, chemical toxins, or radiation damage, represents an inflammatory form of cell death. It is characterized by necrosis, such as cytoplasmic leakage, organelle swelling, plasma membrane rupture, cell lysis, and disintegration. This process results in the release of danger-associated molecular patterns (DAMPs), including cytochrome c, disrupting the intracellular environment and triggering immune activation, ultimately leading to tissue damage [51,52,53]. In contrast to conventional necrosis, necroptosis is governed by distinct regulatory mechanisms.
The necroptotic pathway entails sequential phosphorylation of crucial proteins, specifically receptor-interacting serine/threonine-protein kinase 1 (RIPK1), receptor-interacting serine/threonine-protein kinase 3 (RIPK3), and mixed lineage kinase domain-like pseudokinase (MLKL) [54]. RIPK1, serving as a pivotal initiator, associates with RIPK3 to form and phosphorylate the necrosome. Subsequently, activated RIPK3 phosphorylates MLKL, resulting in its oligomerization and the formation of pores in the plasma membrane. This cascade leads to calcium influx, membrane rupture, and necroptosis [55]. In certain scenarios, necroptosis can occur independently of RIPK1. Z-DNA binding protein 1 (ZBP1), a crucial sensor of necroptosis, can directly bind RIPK3 and MLKL through its RHIM domain, activating RIPK1-independent necroptosis [56]. Therefore, RIPK3/MLKL are central to the execution of necroptosis, independent of caspases [51]. Moreover, necroptosis can be facilitated by the PANoptosome complex, in which ZBP1 and the RIPK3 complex recruit caspase-8, activating the ZBP1-NLRP3 inflammasome and orchestrating the PANoptotic process. This complex is involved in both pyroptosis and apoptosis [57]. In addition to its role in mediating cell death, RIPK3/MLKL also impacts exosome release, as these components are encapsulated in exosomes and transferred to recipient cells [58].
AutophagyAutophagy, a cellular mechanism for self-digestion and degradation, represents a form of programmed cell death. Various stressors, including starvation, radiation, hypoxia, bacterial invasion, and growth factor deficiency, can induce autophagy, which is characterized by cytoplasmic vacuolization and the formation of autophagosomes. After the fusion of autophagic vacuoles and phagosomes, autophagosomes engulf cellular components for subsequent degradation. Microtubules play a crucial role in transporting autophagosomes to lysosomes for degradation. Unlike apoptosis, autophagy is governed by autophagy-related (ATG) genes. These genes selectively target damaged organelles and protein aggregates for lysosomal digestion, preventing cell damage [59].
The initiation of autophagy is catalyzed by the activation of the ULK1 complex, which is composed of ULK1, FIP200, and ATG proteins (ATG12 and ATG101). This complex phosphorylates PI3KC3, resulting in the formation of the ULK1-PI3KC3 complex, subsequently activating PI3P. During this process, additional ATGs, including ATG5, ATG12, ATG16L1, and LC3, are recruited. Simultaneously, LC3-I is phosphorylated, generating lipidated LC3-II on autophagosomal membranes—an indispensable marker of autophagy initiation [60].
Autophagy encompasses macroautophagy, microautophagy, and chaperone-mediated autophagy (CMA) [61]. Macroautophagy entails the sequestration of large cytoplasmic portions in autophagosomes. These structures then fuse with lysosomes to create autolysosomes for subsequent degradation and recycling. Microautophagy directly engulfs cytosolic components through invagination of the lysosomal membrane. CMA depends on Hsc70 to recognize and translocate specific substrate proteins into lysosomes for degradation [62].
In summary, autophagy is an intracellular catabolic process in which autophagosomes encapsulate and degrade harmful or redundant substances into recyclable biomolecules. Autophagy plays a vital role in waste removal, providing energy and nutrients, and regulating cellular differentiation, development, and immunity. Exosomes derived from autophagy, distinct from those released by autophagosomes, transport autophagic signals and bioactive cargo in an ATG-dependent manner [63].
PyroptosisPyroptosis, a form of programmed inflammatory cell death, is characterized by both apoptosis and necrosis. It initiates chromatin condensation, followed by plasma membrane rupture, cell swelling, and lysis [64].
Pyroptosis can be divided into classical and nonclassical pathways. The classical pathway, which is dependent on caspase-1, is triggered by pathogen- or danger-associated molecular patterns (PAMPs/DAMPs). This pathway results in NLRP3 oligomerization, recruitment of pro-caspase-1, and the expression of apoptosis-associated speck-like protein (ASC), ultimately leading to the formation of the NLRP3 inflammasome. Activated caspase-1 cleaves pro-IL-1β/IL-18 into mature cytokines and gasdermin D (GSDMD), generating a pore-forming N-terminal fragment. This fragment integrates into the plasma membrane, forming pores that induce cell swelling, lysis, and release of IL-1β/IL-18 [6, 64]. The nonclassical pathway, initiated by LPS, activates human caspase-4/5 or mouse caspase-11, resulting in GSDMD cleavage and subsequent cytokine release [65,66,67]. Pyroptotic cells release inflammatory DAMPs, including IL-1β, IL-18, HMGB1, ATP, and DNA, intensifying inflammation in neighboring cells [68].
Inflammasome-mediated pyroptosis entails the activation of the NLRP3 inflammasome, leading to cell death and the release of inflammatory exosomes. These exosomes encapsulate molecules that enhance immune responses, inflammation, and phagocytosis [69, 70].
In summary, exosomes that cause programmed cell death exhibit diverse release mechanisms, compositions, and functions. They are produced by specific enzymes or pathways, encapsulating mediators for extracellular transport (Fig. 2). These exosomes modulate recipient cell functions, such as growth, differentiation, apoptosis, and immune activation, through endocytosis, membrane fusion, or receptor interaction [71]. This signaling pathway impacts the immune microenvironment and inflammation, contributing to diseases. For instance, exosomes from apoptotic neurons containing misfolded mutant SOD1 can trigger neuronal death and contribute to amyotrophic lateral sclerosis (ALS) [72]. Intestinal epithelial cells damaged by ischemia/reperfusion release exosomes that promote cortical neuron death [73]. Understanding the various programmed cell death modes and their molecular mechanisms is crucial for gaining insights into diseases and identifying therapeutic targets.
Fig. 2
Exosomes derived from distinct programmed cell death pathways exhibit varied molecular compositions and exert diverse functional impacts. Throughout cellular apoptosis, key initiators such as Caspase 3
Comments (0)