
Remember me
Highly pathogenic influenza A (HPAI) H5N1 viruses infect birds, mammals, and humans, often causing severe disease and high mortality rates. In early 2024, HPAI H5N1 clade 2.3.4.4b viruses began circulating among dairy cows in the state of Texas1, resulting in significant morbidity and substantial reduction in milk production and quality2,3,4. Despite many interventions, these and related viruses have rapidly spread to infect more than 900 farms in 16 US states5. The virus has also spread to other animals and is associated with severe and even lethal disease in wild mammals such as foxes, bears, seals, and sea lions; in domesticated cats and dogs; in farmed mink and foxes; and in other livestock, such as goats6. Thus far, at least 67 human H5N1 infections have occurred in association with this epizootic7. Concern is mounting that the epizootic will not be contained with traditional farm biosecurity interventions alone and cattle vaccines against H5N1 are urgently needed.
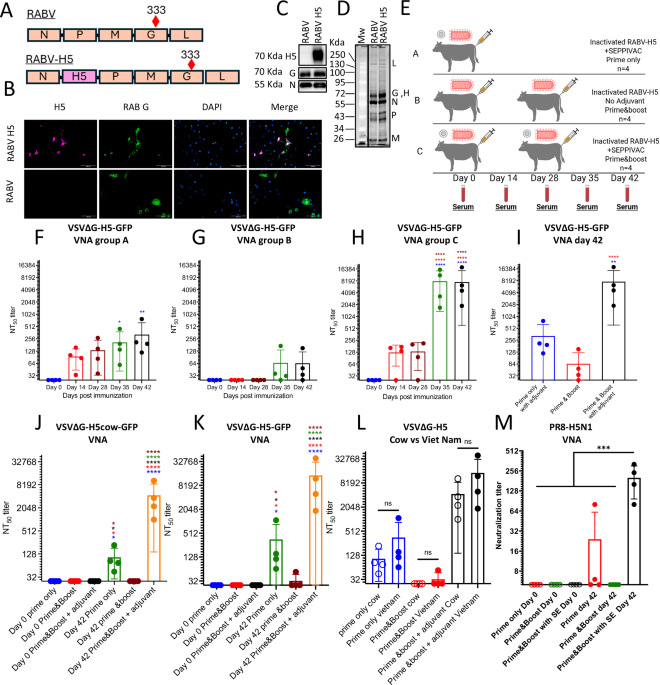
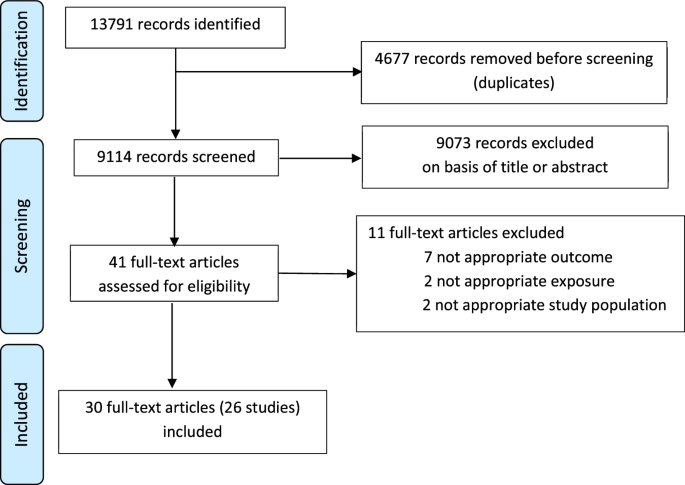
We developed a rabies virus-based H5 inactivated vaccine to address this concern and evaluated its immunogenicity in cattle. To express the H5N1 HPAI HA antigen by the rabies vector, a synthetic full-length codon-optimized HA ORF of the Influenza virus A/Viet Nam 1203/2004(H5N1) isolate was cloned into the well-established rabies vaccine vector (BNSP333)8 between the N and P genes (Fig. 1A). The virus was entitled RABV-H5. Based on previous immunogenicity data in mice (our unpublished data), we used a vaccine harboring the clade 1 Viet Nam 1203/2004 H5 gene. We confirmed the expression of the H5 and rabies G proteins in RABV-H5-infected cells by immunofluorescence with RABV G and H5 specific antibodies (Fig. 1B). The incorporation of H5 in addition to the other structural rabies virus proteins (L, G, N, P and M) was shown by analysis of sucrose-cushion-purified vaccine-virus particles by both SYPRO®-Ruby staining and by western blot analysis (Fig. 1C, D respectively).
Fig. 1: H5 immunogenicity of RABV-H5 vaccine in cattle.
A Schematic genomic maps of the RABV-H5 vaccine vector and the parental RABV. The R333E-attenuating mutation in the RABV G antigen is indicated. N=nucleoprotein, P=Phosphoprotein, M=matrix protein, G=glycoprotein, L=polymerase. B Immunofluorescent staining of H5 and RABG in RABV-H5 and RABV-infected Vero cells. Cells were infected at MOI 0.01 and fixed after 48 h. Cells were permeabilized and stained with α-HPAI-H5N1 H5 polyclonal antibodies (purple), α-RABV-G 4C12 monoclonal antibody (green), and DAPI to visualize the nuclei (blue). In the merged images, white indicates an overlap between purple and green. Images were taken at 40X magnification. Scale bars represent 100 μm. C, D represent protein composition analysis of 1 μg sucrose purified virions by western blotting (C) and SDS-PAGE followed by SYPRO®-RUBY protein staining (D). Blots were either probed with α- H5 (NR-163), α-RABV-G 4C12 (middle panel) or α-RABV-N polyclonal rabbit sera (bottom panel). E Cattle immunization and blood draw schedule; 3 groups of 4 cattle were immunized once (group A) or twice (B, C) with 100 μg/dose of BPL-inactivated RABV-H5 vaccine, with (A, C) or without (B) SEPPIC-SWETM adjuvant. Created with Biorender.com. VSV∆G-H5-GFP virus neutralization in prime only with adjuvant (F), prime-boost without adjuvant (G) and prime-boost with adjuvant (H). Filled circles are NT50 titers. Average (bars) and STDEV (error bars) are shown. Comparison of day 42 NT50 values between the three vaccination groups is shown (I). Day 0 and 42 VNA titers (NT50) against VSV∆G-H5-GFP harboring the H5-cow version (J) or the H5- A/Viet Nam/1203/2004 version (K) and the comparison between them (L). PR8-H5N1 (M) and HPAI-H5N1 cow (N) VNAs (100% neutralization) are shown. Ordinary one-way ANOVA with Tukey’s Multiple Comparison Test was used to determine statistical differences between groups at each time point. Stars indicate significant differences. Error bars indicate the mean with SD for groups of 4 cattle with samples run in duplicate. (****p < 0.0001; ***p = 0.0001; **p ≤ 0.0029; *p < 0.0173; ns not significant).
To assess the immunogenicity of the BNSP333-H5 vaccine in cattle, 12 heifer calves were randomly assigned to 3 vaccination groups: A- received the RABV-H5 vaccine with SEPPIC adjuvant in a prime-only regimen; B- received the vaccine without adjuvant in a prime-boost regimen; and C- received the vaccine with SEPPIC adjuvant in a prime-boost regimen (Fig. 1E). The RABV-H5 vaccine was Betapropiolactone (BPL) inactivated and a 100 µg was injected subcutaneously (SC). The 100 µg vaccine dose was selected based on previous immunogenicity studies using the RABV-based vaccine in non-human primates9. The SC delivery is commonly used for beef cattle vaccinations (e.g. respiratory and clostridial vaccines that are part of the vaccination program for cattle and calves on cow-calf operations in the US). The SC route is also preferred to avoid affecting carcass quality. The cattle were not euthanized, and anesthesia before vaccine delivery or blood draws is not required for large animal species.
Serum from all vaccinated animals was collected on days 0, 14, 28, 35 and 42 post-prime for virus-neutralizing antibodies (VNA). Cattle studies were completed at the USDA-ARS, Livestock Arthropod Pest Research Unit (Knipling-Bushland US Livestock Insects Research Laboratory (KBUSLIRL), Kerrville, TX, USA), and all animal procedures were approved by the KBUSLIRL Institutional Animal Care and Use Committee (IACUC) (Protocol #2024-05).
To analyze the vaccine-induced VNA against HPAI H5N1, we used a recombinant vesicular stomatitis virus (VSV) in which the VSV-G surface glycoprotein was deleted and replaced with HPAI H5 (Clade 1, A/Viet Nam/1203/2004(H5N1) with the intact polybasic cleavage site. In addition, the gene-encoding green fluorescent protein (GFP) was also incorporated into the genome, to generate VSV∆G-H5-GFP.
Our results presented in Fig. 1F indicate neutralizing antibodies to H5 14 days after a single immunization by the adjuvanted vaccine, and the neutralization titer was maintained throughout the study. A booster immunization with adjuvant on day 28 significantly increased the VNA titer (Fig. 1H). The unadjuvanted vaccine did not induce detectable VNAs after the prime and low VNA were detected after the boost immunization (Fig. 1G). At day 42 the highest VNA titers were measured in sera of prime-boost-immunized animals with the adjuvanted vaccine, whereas all animals that received the single immunization demonstrated a significant but lower VNA response, indicating successful priming (Fig. 1I). Prime-boost immunization without adjuvant showed VNA only in one out of four animals.
Having demonstrated that the RABV-H5 vaccine was capable of inducing H5-VNA, we asked whether the vaccine that carries a clade 1 H5 (A/Viet Nam/1203/2004(H5N1)) would induce cross VNA to the currently circulating 2.3.4.4b clade. Thus, using the above-mentioned recombinant VSV, we replaced the clade 1 H5 with the H5 gene of an HPAI from the currently circulating 2.3.4.4b clade. Indeed, sera from cattle vaccinated with the vaccine encoding the H5 clade 1 related antigen (A/Viet Nam/1203/2004(H5N1) efficiently neutralized VSV∆G-H5cow-GFP representing the circulating antigen (Clade 2.3.4.4b Cow) in both group A (Prime only with adjuvant) and group C (Prime-boost with adjuvant) (Fig. 1J). Testing the neutralization capacity of the clade 1 H5 (A/Viet Nam/1203/2004(H5N1) with parallel serum dilution revealed a similar pattern of neutralization, namely that sera from both groups A and C efficiently neutralized the VSV∆G-H5 Vietnam-GFP (Fig. 1K). Of note, the VNA titers towards clade 1 or clade 2.3.4.4b were not significantly different (Fig. 1L), indicating the potential of the RABV-H5 vaccine candidate to confer cross-clade protective immunity.
To confirm the results detected with the VSV∆G-H5-GFP, we determined the neutralizing antibody titers against PR8-H5N1, a recombinant Puerto-Rico 8 influenza A virus in which the HA and NA genomic segments have been replaced with the respective segments of H5N1(A/Viet Nam/1203/2004(H5N1) and the polybasic cleavage site has been deleted as previously reported10. Similar to the neutralization results obtained with VSV∆G-H5-GFP, a prime-boost vaccination with an adjuvanted vaccine (group C) induced significantly higher neutralizing antibodies compared to other vaccination regimens or to day 0 serum samples (Fig. 1M).
Analysis of the serum samples for rabies-virus-neutralizing titers revealed that already 14 days post single immunization with an adjuvanted vaccine, rabies-virus-neutralizing titers were above 0.5 IU/mL, the WHO-accepted protective antibody level against RABV (Fig. 2A). The rabies-virus-neutralizing titer remained significantly high on day 28 and then retreated but remained above the WHO-accepted protective antibody level (0.5 IU/ml) (Fig. 2A). Similarly to H5 neutralization, rabies neutralizing titers were significantly higher following prime-boost vaccination with adjuvant (Fig. 2C). Unadjuvanted vaccine required two immunizations to induce low, yet significant, neutralizing antibody response (Fig. 2B). Comparing the responses of the three vaccination groups on day 42 revealed that prime-boost with adjuvant (group C) yielded significantly higher titers than groups A and B (Fig. 2D), which was similar to the neutralization of both VSV∆G-H5-GFP (Fig. 1I) and PR-8 H5N1 viruses (Fig. 1M, N).
Fig. 2: Vaccine immunogenicity in the presence and absence of preexisting RABV immunity.
Rabies virus (strain CVS-11) VNAs (IU/ml) in cattle sera were determined by Rapid fluorescent focus inhibition test (RFFIT) in prime only with adjuvant (A), prime- boost without adjuvant (B) and prime-boost with adjuvant (C). Comparison of day 42 VNA titers is shown (D). Cattle immunization and blood draw schedule (E). Six cattle were immunized twice with 100 μg/dose of BPL-inactivated, SEPPIC-SWETM adjuvanted RABV-GP38 vaccine. On Day 200, the cattle were vaccinated with the same dose of RABV-H5 with SEPPIC-SWE; and on day 2283 cattle received another dose of the adjuvanted RABV-H5 vaccine. Syringes represent immunizations, and blood tubes indicate the days blood was drawn. Created with Biorender.com. VSV∆G-H5-GFP virus-neutralization titers (NT50) (F), PR8-H5N1 virus-neutralization titers (100% neutralization) (G), A/cattle/Texas/56283/2024 (H5N1) virus-neutralization titers (100% neutralization) (H) and Rabies virus-neutralization titers (IU/ml) (I) are shown. Open circles in (I) refer to cows receiving 1 dose of the H5N1 vaccine 3 total doses of the RABV vector while closed circles refer to 2 H5 doses and 4 total RABV doses. Bars represent the mean values, and error bars indicate standard deviation. Ordinary one-way ANOVA with Tukey’s Multiple Comparison Test was used to determine statistical differences between groups (****p < 0.0001; ***p ≤ 0.0008; **p ≤ 0.0024; *p ≤ 0.047; ns not significant). The dotted line indicates 0.5IU/mL, the WHO-accepted protective antibody level against RABV.
The safety profile of the inactivated rabies vaccine platform is well-established based on multiple studies and is currently entering a phase 1 clinical trial (ClinicalTrials.gov NCT 06546709). We used a commercially available adjuvant that is approved for use in both animals and humans (SEPIVAC SWE™) and a thorough analysis of blood samples from the vaccinated cattle revealed biochemical and hematological values as expected in healthy animals substantiating the safety profile of both our vaccine and the SEPIVAC SWETM adjuvant (Supplementary Figs. 1–5).
Moreover, we wanted to determine the immunogenicity of the RABV-H5 vaccine in the presence of preexisting RABV immunity. Using the SEPIVAC-SWETM adjuvanted rabies-H5 vaccine, we vaccinated cattle (n = 6) at 200 days after they had been immunized (prime-boost with SEPIVAC-SWETM) with an unrelated RABV vaccine from another study (RABV-GP3811, Fig. 2E). Blood was collected as indicated and analyzed for H5 and RABV-neutralizing antibodies (Fig. 2E).
Indeed, we found that cattle previously vaccinated with the RABV-H5 vaccine mounted a robust VSV∆G-H5-GFP and PR8-H5N1 neutralization response (Fig. 2F, G, respectively), similar to the response of vaccinated animals without preexisting immunity to the RABV vaccine vector (Fig. 1H, M, respectively). For neutralization of both VSV∆G-H5-GFP and PR8-H5N1 viruses, a prime-boost of the H5 vaccine was significantly better than a prime-only regimen (Fig. 2F, G). We further tested the capacity of the same cattle sera to neutralize the circulating HPAI virus (A/cattle/Texas/56283/2024 (H5N1))12. As early as 14 days after the first H5 vaccination (day 214) one animal already had detectable neutralizing antibodies. By day 42 post H5 vaccination (day 242), all prime-boost vaccinated animals and one animal that received only a single dose (“prime only”) had neutralizing antibodies (Fig. 2H). Importantly, the day 242 neutralizing titers following a prime-boost regimen were not significantly different from the titers of naturally H5N1 infected and convalescent cows, yet additional samples of convalescent sera are needed to substantiate these results. Analysis of rabies-virus-neutralizing antibodies revealed that after 200 days, rabies-neutralizing titers were lower than day 42, yet were above the WHO protective value (0.5 IU/ml). Despite preexisting immunity to rabies, vaccination with an adjuvanted rabies-H5 vaccine induced a robust rabies-virus-neutralization immune response (Fig. 2I), similar to the H5 VNAs (Fig. 2F, G). A single boost (third dose of the RABV antigens) on day 200 was sufficient to induce robust RABV VNAs despite preexisting immunity, and an additional vaccine dose (fourth dose) had no significant benefit (Fig. 2I, compare open and closed circles).
The ability of vaccines to confer cross clade protection against the circulating HPAI-H5N1 is important. While mouse study by Hawman and collaborators13 using a replicating RNA vaccine indicated that a clade 2.3.4.4b, but not clade 1 HA-based vaccine, protects against H5N1 (cattle) challenge in mice, our cattle study and importantly human studies demonstrated the ability of clade 1 H5N1 vaccine to mount clade 2 immunity. Others demonstrated the induction of clade 2.3.4.4b-cow antibodies (neutralizing and acting through ADCC) in humans vaccinated with an A/Viet Nam/1203/2004(H5N1) vaccine14,15. Additionally, the observed milder disease in aged immune individuals and the protective role of vaccine history in animal models16,17 further support our results, demonstrating cross-clade virus neutralization in immunized cattle.
In the current study and in light with previous publications18,19, we show that using VSV∆G-H5-GFP (clade 1 and clade 2.3.4.4b) for virus neutralization assays (Figs. 1 and 2) gave a similar pattern of neutralization when using the same samples with either PR8-H5N1 or HPAI-H5N1 (A/cattle/Texas/56283/2024). Besides ease of performance for multiple samples and no safety concerns, pseudoviruses allow for precise evaluation of VNAs towards H5, reducing the possible responses to other influenza viruses from previous natural exposures. Despite the differences between the different pseudoviruses used here and in previous studies18,19, and the fact that we applied cattle rather than human sera, we similarly observed higher neutralization titers with the recombinant VSV. Unravelling the reasons for these differences is however beyond the scope of this manuscript.
Our study has several limitations. First, beef cattle were used instead of dairy cows. Although the immune response should also be tested in milk-producing cows, we believe that this cattle vaccination study supports the further development of the RABV-H5 vaccine to control the virus spread and mitigate the pandemic risk. Second, to expedite the study, we focused on neutralizing antibody titers and did not study other mechanisms of protection such as ADCC. Third, we could not include a virus challenge step in the current study.
We show that the vaccine induced VNAs against HPAI-H5N1 (A/cattle/Texas/56283/2024) were not significantly different from convalescent samples (Fig. 2H). Neutralizing antibodies play an important role in protection against influenza in general and particularly against HPAI-H5N120,21,22. Unfortunately, correlates of protection in the serum of cattle against the circulating clade are not known. Convalescence following infections confer at least partial protection from infection with the homologous clade20. Demonstrating that VNAs in vaccinated and convalescent sera were not significantly different, allows us to suggest that the vaccine induces protective neutralizing antibodies titers.
In summary, we demonstrated the safety and immunogenicity of this RABV-H5 vaccine in cattle. Despite the limitations of this study, the increasing awareness of the virus spillover to other animal species and humans warrants protective measures. The RABV vaccine platform has demonstrated long-term performance in multiple hosts, including mice, hamsters, nonhuman primates, and humans. Having a rabies based HPAI-H5N1 dual vaccine is valuable in certain world regions where rabies is endemic and transmitted to cattle from bats23,24. This further aligns with the one health approach to protect animals and humans from zoonotic infections (rabies and HPAI-H5N1). Thus, we believe the results obtained with the rabies-H5 vaccine candidate support its further development as a vaccine candidate for HPAI H5N1.
Comments (0)