
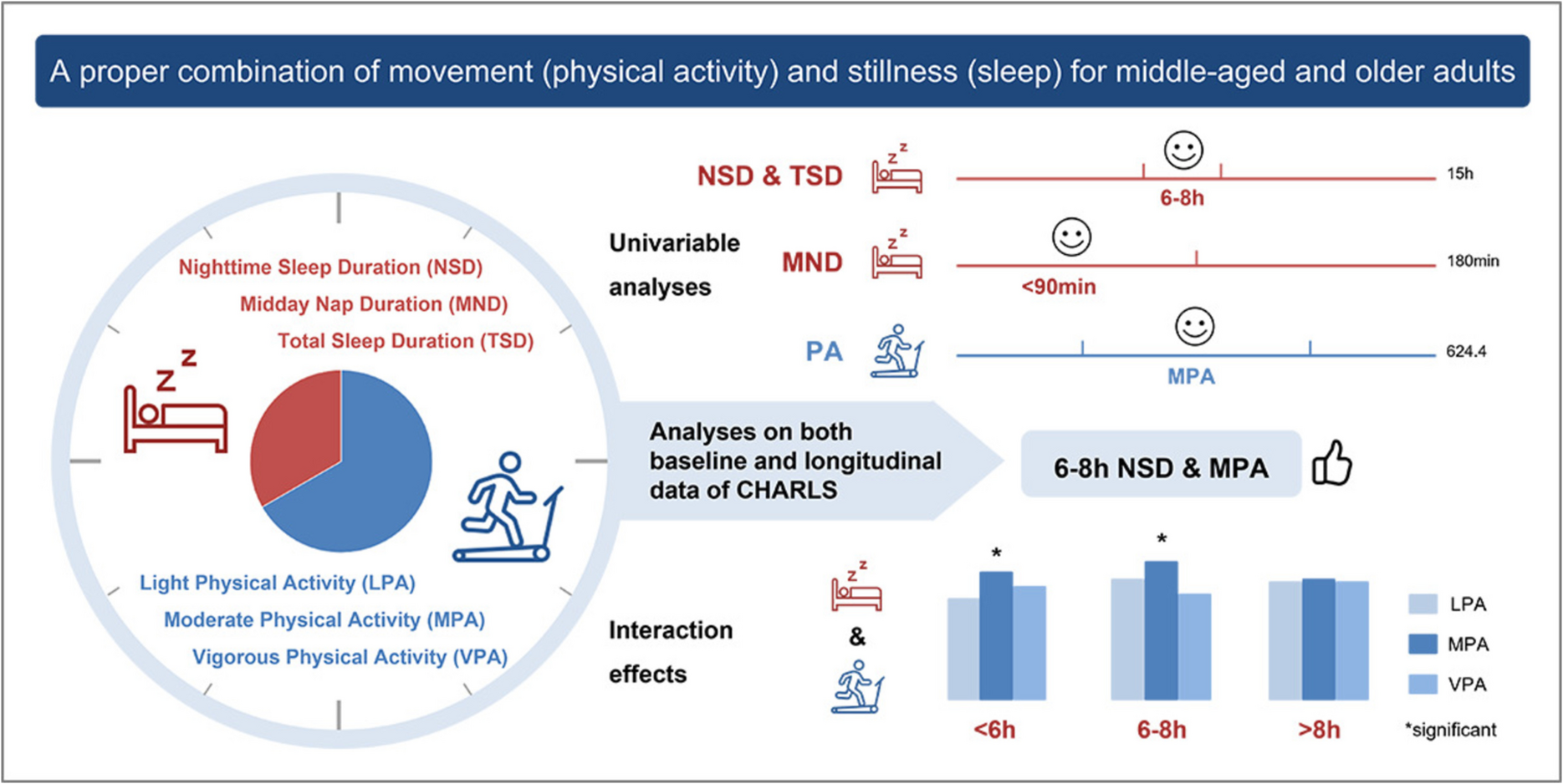
Remember me



In older adults, higher-intensity exercises improve endurance capacity more than moderate-intensity exercises [32, 33]. To assess whether this holds in older rats, we compared the effects of standardized MICT and HIIT on endurance performance using an incremental exercise test at baseline (PRE) and after a 4-week training (POST) to measure SLT and Smax(Fig. S1). Repeated measures ANOVA revealed a significant main Time effect for both SLT (F(1,16) = 78.92, p < 0.0001) and Smax (F(1,16) = 167.4, p < 0.0001). Additionally, a significant Time × Group interaction was observed for SLT (F(2,16) = 15.96, p < 0.001) and Smax (F(2,16) = 39.56, p < 0.001), suggesting that the magnitude of improvement varied between training groups. Sidak post hoc analysis revealed that within trained groups SLT and Smax values were higher in POST than in PRE (p < 0.0001). Regarding the differences between groups, Tukey post hoc analysis suggested that SLT and Smax values were higher in both MICT (p < 0.01) and HIIT (p < 0.001) compared to CONTROL in POST (Fig. S1B and C). HIIT group had greater Smax compared to MICT in POST (p < 0.05) (Fig. S1C).
MICT had greater effect on hippocampal and cortical energy metabolism-related protein expressionBecause the efficacy of an endurance exercise program is highly dependent on the dosage between the intensity and the duration, we sought to identify the molecular signature in the cerebral cortex and hippocampus related to energy metabolism specific to HIIT and MICT. We therefore removed these brain regions at 26–30 h after the last exercise test for all molecular analyses.
In the cortexFirst, we measured the levels of AMPKα, as well as COX4, CS, and a key metabolic regulator that interacts with brain-derived neurotrophic factor (BDNF), the estrogen-related receptor alpha (ERRα) [16]. Welch ANOVA analysis revealed a significant group effect for COX4 (F(2,5.76) = 8.95, p < 0.05), and pAMPKα/AMPKα ratio F(2,7.05) = 8.48, p < 0.05), while one-way ANOVA revealed significant group effect for ERRα (F(2,14) = 8.08, η2 = 0.54, p < 0.01) and CS (F(2,16) = 22.77, η2 = 0.74, p < 0.0001). Dunnett’s post hoc revealed that HIIT had a higher pAMPKα/AMPKα ratio (p < 0.05) and also tended to have higher COX4 levels (p = 0.06) relative to CONTROL (Fig. 1A, B). Tukey post hoc showed higher ERRα in MICT compared to HIIT and CONTROL (p < 0.05) while CS was upregulated in both MICT and HIIT compared to CONTROL (p < 0.001) (Fig. 1C, D). In addition, we investigated the expression of the cerebral lactate transporters following MICT and HIIT. This major energy metabolite is known to promote the expression of neurotrophic factors, including BDNF and VEGF, and may thus mediate cerebral plasticity [21]. One-way ANOVA revealed significant differences between groups for cortical MCT2 (F(2,16) = 4.39, η2 = 0.35, p < 0.05) and MCT4 (F(2,14) = 4.97, η2 = 0.42, p < 0.05) suggesting that lactate transporters were sensitive to MICT. Indeed, Tukey post hoc demonstrated that MICT had higher MCT2 levels compared to CONTROL (p < 0.05), whereas MCT4 expression was lower in HIIT relative to MICT (p < 0.05) (Fig. 1E, F).
Fig. 1
Effect of training regimens on cortical proteins involved in metabolic activity by western blot. Level of cortical A pAMPKα/AMPKα ratio (n = 19), B COX4 (n = 18), C ERRα (n = 17), D CS (n = 19), E MCT2 (n = 19), F MCT4 (n = 17), G VEGF (n = 19), and H VEGFR-2 (n = 17) in MICT, HIIT, and CONTROL groups at POST. Pictures of the western blot membranes are shown below each graph. *Significant differences in protein levels between groups. Data is expressed in mean ± SD
We then examined the influence of endurance training on protein markers of angiogenesis. One-way ANOVA showed significant differences between groups for VEGF (F(2,16) = 4.44, η2 = 0.36, p < 0.05) and its receptor VEGFR-2 (F(2,14) = 9.58, η2 = 0.58, p < 0.01). Indeed, VEGF levels were higher in MICT compared to CONTROL, while MICT upregulated VEGFR-2 compared to the two other groups (Tukey post hoc: p < 0.05 for all) (Fig. 1G, H). No statistical difference between groups was found for AMPKα, pAMPKα, PGC-1α, MFN1, MFN2, MCT1, PECAM-1, the N-methyl-D-aspartate Receptor-1 (GluN1), BDNF, and CREB (Fig. S4 and S6).
In the hippocampusIn accordance with studies showing that endurance training improved mitochondrial activity [34], one-way ANOVA demonstrated significant differences between groups for AMPKα (F(2,16) = 3.75, η2 = 0.32, p < 0.05), pAMPKα (F(2,16) = 4.91, η2 = 0.38, p < 0.05), pAMPKα/AMPKα ratio (F(2,16) = 4.32, η2 = 0.35, p < 0.05), COX4 (F(2,16) = 4.65, η2 = 0.38, p < 0.05), CS (F(2,16) = 6.2, η2 = 0.44, p < 0.05), and for the mitofusin proteins, i.e., MFN1 (F(2,16) = 14.13, η2 = 0.64, p < 0.001) and MFN2 (F(2,16) = 3.82, η2 = 0.32, p < 0.05). Tukey post hoc showed that MICT upregulated AMPKα, pAMPKα, pAMPKα/AMPKα ratio, CS, and COX4 levels compared to CONTROL (p < 0.05), while COX4 also tended to be higher in HIIT compared to CONTROL (p = 0.06) (Fig. 2A, B, C, D, E). In contrast, while MFN2, protein involved in mitochondrial fusion level, was only higher in HIIT compared to CONTROL (p < 0.05), HIIT had higher MFN1 level compared to the two other groups (p < 0.01, Tukey post hoc for all) (Fig. 2F, G). In the same way, we quantified the master regulator of mitochondrial biogenesis, namely PGC-1α, which is also involved in cerebral plasticity processes by upregulating BDNF. One-way ANOVA revealed significant differences between groups (F(2,16) = 6.61, η2 = 0.45, p < 0.01). Higher levels were found in MICT compared to CONTROL (Tukey post hoc: p < 0.01) (Fig. 2H).
Fig. 2
Effect of training regimens on hippocampal proteins involved in metabolic activity by western blot. Level of hippocampal A AMPKα (n = 19), B pAMPKα (n = 19), C pAMPKα/AMPKα ratio (n = 19), D CS (n = 19), E COX4 (n = 19), F MFN1 (n = 19), G MFN2 (n = 19), H PGC-1α (n = 19), I VEGF (n = 18), J VEGFR-2 (n = 19), K MCT4 (n = 17), and L GluN1 (n = 19) in MICT, HIIT, and CONTROL groups at POST. Pictures of the western blot membranes are shown below each graph. *Significant differences in protein levels between groups. Data is expressed in mean ± SD (A–K) and in median (min to max) (L)
Significant differences between groups were also found for angiogenesis and metabolic activity markers such as VEGF (Welch ANOVA: F(2,6.86) = 5.4, p < 0.05) and VEGFR-2 (one-way ANOVA: F(2,16) = 3.72, η2 = 0.32, p < 0.05). Higher VEGF (Dunnett’s post hoc: p < 0.05) were found in the MICT compared to CONTROL (Fig. 2I), while MICT had also higher levels of VEGFR-2 compared to HIIT (Tukey post hoc: p < 0.05) (Fig. 2J). Endurance training induced changes in the hippocampal lactate cotransporters MCT4 as shown by significant difference between groups (one-way ANOVA: F(2,14) = 4.46, η2 = 0.39, p < 0.05). HIIT downregulated MCT4 expression compared to MICT (Tukey post hoc: p < 0.05) (Fig. 2K). Finally, significant differences were found between groups for GluN1, which can be considered as a synaptic plasticity marker (Kruskal Wallis test: (H(2) = 8.979, p < 0.01). Dunn’s post hoc suggested that MICT had higher GluN1 level as compared to CONTROL (p < 0.01). There was no statistical difference between groups for ERRα, MCT1, MCT2, PECAM-1, CREB, and BDNF (Fig. S5 and S6). No difference was found for α-tubulin normalized by GAPDH and Ponceau staining (Fig. S7).
MICT induced more extensive transcriptomic changes compared to HIIT Bulk RNA-seq in the cortexCompared with CONTROL, differentially expressed genes (DEGs) analysis (p < 0.05) of MICT and HIIT revealed 235 and 38 DEGs respectively. Of these, 160 and 22 genes were upregulated, while 75 and 16 genes were downregulated in MICT and HIIT groups, respectively. While 27 DEGs were identified as common to both exercise conditions, MICT specifically upregulated 139 genes and downregulated 69 genes, while HIIT only increased the expression of one gene and reduced the expression of ten genes (Fig. 3A).
Fig. 3
Both training regimens influence metabolic activity and plasticity-associated transcriptional landscape in the cortex. A Venn diagram representing the number of differentially expressed genes (DEGs) in both MICT and HIIT groups compared to CONTROL from a bulk RNA sequencing of cortical brain tissue (n = 4–8 rats per group). The distribution of genes that were either up- or down-regulated is also shown. Percentages were calculated from the total upregulated and downregulated DEGs (MICT and HIIT). B Global DEGs in MICT compared to CONTROL. C Global DEGs in HIIT compared to CONTROL
For example, MICT increased the expression of Didt4, Nfkbia, Sgk1, Gpd1, Olig2, Pdk4, Nr4a3, and Fam107a, while reducing that of Ier2 (Fig. S8A). DEGs were associated with various GO terms/pathways including developmental-like processes (e.g., cell differentiation and proliferation), rhythmic process, and metabolic processes (Fig. 3B). Similarly, the genes that were upregulated with MICT were involved in the aforementioned functions (Fig. S8C).
Among others, HIIT strongly increased the expression of Nfkbia, Didt4, Sgk1, Gpd1, Olig2, and Tagap while reducing the expression of Ier2 and Tbx1, involved in different metabolic processes that were observed with MICT (Fig. 3C, S8B, and S8D). While the metabolic process was identified as common to both exercise conditions, MICT influenced specific biological functions also involved in brain development processes (Fig. S11A and B).
To gain further insight into the mechanisms of action of MICT and HIIT, the Ingenuity Pathway Analysis regulator (IPA) was used to predict molecules that may be responsible for changes in gene expression. This tool predicted that MICT activated various upstream regulators including the CREB pathway and the 8 bromo cAMP (which activates the cyclic AMP-dependent protein kinase involved in glucose and lipid metabolism, gene transcription, and cell growth). Furthermore, the IPA predicted the activation of the ERK pathway, which is mediated both by the activation of pathways related to tetradecanoylphorbol acetate and the inhibition of pathways triggered by the ERK/MEK pathway inhibitor PD98059. HIIT was predicted to inhibit fewer upstream regulators activated by MICT, including the dexamethasone phosphate and the dexamethasone (anti-inflammatory effect) dependent pathways (Fig. S9).
Bulk RNA-seq in the hippocampusCompared with CONTROL, 57 and 29 DEGs were found in MICT and HIIT, respectively (p < 0.05). Among them, 31 and 7 genes were upregulated, while 26 and 22 genes were downregulated with MICT and HIIT, respectively. While 27 DEGs were identified as common to both exercise conditions, MICT specifically upregulated 26 genes while downregulating 19 genes. HIIT specifically upregulated only two genes and downregulated 15 genes (Fig. 4A).
Fig. 4
Both training regimens influence metabolic activity and plasticity-associated transcriptional landscape in the hippocampus. A Venn diagram representing the number of differentially expressed genes (DEGs) in both MICT and HIIT compared to CONTROL from a bulk RNA sequencing of hippocampal brain tissue (n = 4–8 rats per group). The distribution of genes that were either up- or down-regulated is also shown. Percentage was calculated from the total upregulated DEGs (MICT and HIIT) and downregulated DEGs (MICT and HIIT). Volcano plot representing all the genes Gene Ontology (GO) enrichment/pathways analysis of B global DEGs and C up-regulated DEGs in MICT compared to CONTROL. D GO analysis of global DEGs in HIIT compared to CONTROL
MICT mainly increased the expression of Didt4, Mertk, Fam107a, Btg2, and Rbm3 genes, which showed enrichment of GO terms/pathways involved in metabolic and neuronal functions (Fig. 4B and S10A). More specifically, most genes upregulated with MICT were implicated in the regulation of metabolic activity and the Notch signaling pathway (Fig. 4C). The latter is known to play a role in both neurogenic and synaptic plasticity processes.
HIIT mainly increased the expression of Lfng and Tp53bp2 and decreased the expression of Adamts9. The corresponding GO terms/pathways included different metabolic pathways than MICT (Fig. 4D and S10B). While the Notch signaling pathway was identified as being common to both exercise conditions, MICT specifically regulated genes expression involved in neuronal energy metabolism and differentiation (Fig. S11C).
The IPA predicted that MICT would stimulate the dexamethasone-activated pathways that have anti-inflammatory effects. For the HIIT, no activation/inhibition prediction was found in the hippocampus (Fig. S10C). The primary networks predicted to be involved in metabolic and neurotrophic activity in the cortex and hippocampus are presented in figs. S12, S13, S14, S15. These figures suggested that MICT promotes stable metabolic adaptations and neurogenesis with reduced stress, while HIIT induces stronger plasticity but requires repair mechanisms, particularly in the hippocampus.
Endurance performance is correlated with metabolic activity markers In the cortexOur results showed positive correlation between endurance and some metabolic activity markers. Indeed, SLT was positively correlated with VEGF (r = 0.55, large effect size, p < 0.05), MCT2 (r = 0.61, large effect size, p < 0.01), CS (r = 0.61, large effect size, p < 0.01), and COX4 (r = 0.63, large effect size, p < 0.01). In the same way, positive correlations were found between Smax and MCT2 (r = 0.66, large effect size, p < 0.01), CS (r = 0.61, large effect size, p < 0.01), and COX4 (r = 0.65, large effect size, p < 0.01) (Fig. S16).
In the hippocampusSLT was positively associated with MFN1 (r = 0.61, large effect size, p < 0.01), MFN2 (r = 0.58, large effect size, p < 0.05), and COX4 (r = 0.66, large effect size, p < 0.01). Furthermore, positive correlations were found between Smax and MFN1 (r = 0.61, large effect size, p < 0.01) and COX4 (r = 0.65, large effect size, p < 0.01) (Fig. S17).
Comments (0)