
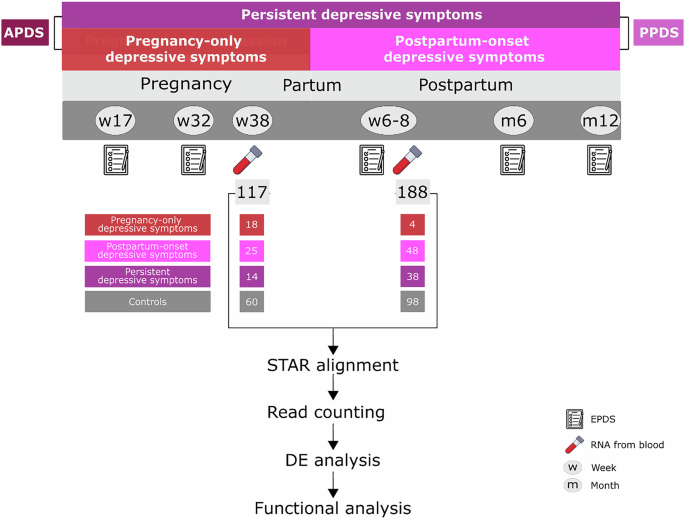
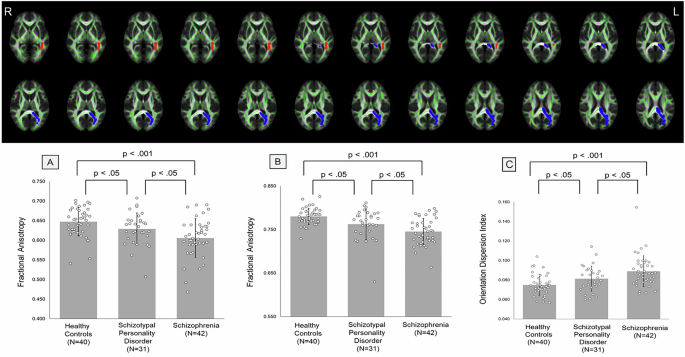
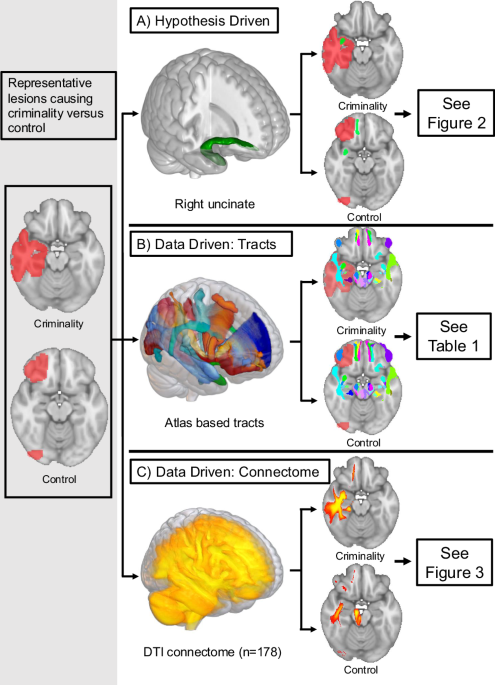
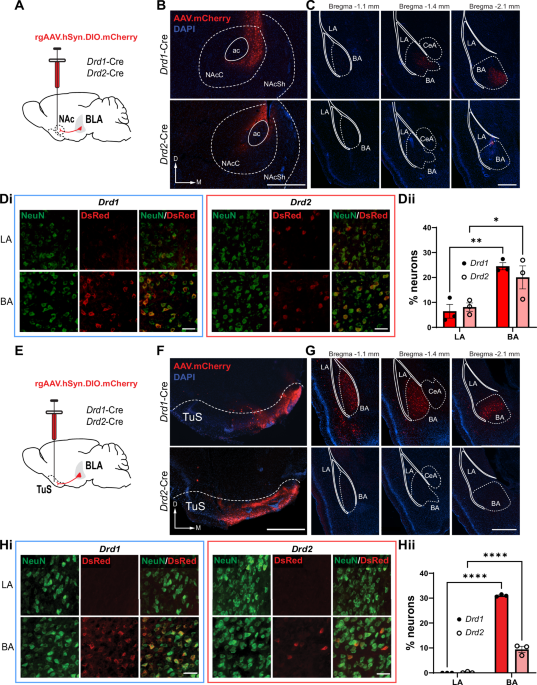
Remember me


We collected clinical, brain MRI, and genetic data from 14 FOXG1 syndrome cases, with 5 from our hospital and 9 from the literature [17, 18, 39, 40]. All cases carried intragenic heterozygous FOXG1 variants, including p.Gln86AspfsTer34 (Q86Dfs*34), p.Gln86ArgfsTer106 (Q86Rfs*106), p.Glu136Ter (E136X), p.Pro182Gln (P182Q), p.Asn187Lys (N187K), p.Ile194Ser (I194S), p.Phe215Leu (F215L), p.Arg230His (R230H), p.Leu235His (L235H), p.Gly252Val (G252V), p.Gly252Asp (G252D), p.Trp255LeufsTer156 (W255Lfs*156), p.Tyr307Ter (Y307X), and p.Tyr416Ter (Y416X) (Table 1). These variants covered different FOXG1 domains (Fig. 1A), including 3 N-terminal early terminations (2 frameshift and 1 nonsense), 9 variants within or near the FBD and CS domains (8 missense and 1 frameshift), and 2 C-terminal nonsense variants (Table 1, Fig. 1A).
Table 1 Genetic and phenotypic features of individuals with FOXG1 syndrome.Fig. 1: Variant locations in FOXG1 and their expression levels.
A Localization of the 14 FOXG1 variants analyzed in this study. Three variants lead to early termination at the N terminus, and three variants result in truncation at the C terminus, while the remaining 8 variants are missense variants within or in proximity to the CS and FBD domains. Variants causing different severities of brain anomalies are color-coded: Red for severe, green for moderate, blue for mild, and black for normal. B Protein expression of FOXG1 and its variants in transfected cells, determined by western blotting with Myc antibody. GAPDH served as the loading control. C Bar graph showing the relative expression of FOXG1 variants compared to WT FOXG1 in ascending order of expression (n = 3 independent transfections). D Bar graph presenting the relative expression of FOXG1 variants on a log2 scale (n = 3 independent transfections). Error bars: S.E.M. *p < 0.05, **p < 0.01, ****p < 0.0001. One-way ANOVA; post-hoc: Uncorrected Fisher’s LSD.
The predominant clinical manifestation was microcephaly, affecting 13 out of 14 cases. A comprehensive analysis of brain MRIs (Table 1, Supplementary Fig. 1) revealed distinct neuroimaging features, with anomalies in the corpus callosum (11/14) and a simplified gyral pattern (11/14) being the most prevalent. This was followed by hypoplasia of the frontal lobes (9/14), enlargement of inner CSF spaces (8/14), and hypoplasia of the basal ganglia relative to the thalami (7/14). Anomalies within the corpus callosum were further categorized as partial agenesis in 7 individuals and thinning in 4 individuals.
Clinical severity ranged from mild to severe, with N-terminal variants presenting moderate to severe clinical severity, while those with pathogenic variants within the FBD domain exhibited heterogeneous severity, ranging from mild to severe. Individuals with variants within the C-terminal region displayed moderate clinical severity. The brain MRI severity score, reflecting the summation of brain anomalies, also ranged from mild to severe (Table 1). Interestingly, N-terminal variants correlated with severe brain MRI anomalies, whereas FBD domain variants exhibited heterogeneous severity. Variants in the C-terminal region were linked to moderate-to-severe brain anomalies. Although there was some correlation between variant location and clinical features, they did not provide a perfect prediction of severity.
Low-expressing Foxg1 variants are linked to moderate-to-severe brain anomaliesTo investigate the effects of different variants on FOXG1 expression, we cloned and transfected Myc-tagged Foxg1 (pCMV-Myc-Foxg1) into U87-MG human glioblastoma cells. The FOXG1 protein localized correctly within the nucleus (Supplementary Fig. 2). Subsequently, disease-associated variants were generated and transfected into U87-MG cells, and protein expression was analyzed after two days in culture (DIV) by western blotting (Fig. 1B). Among the nonsense variants, E136X exhibited very low protein expression, whereas Y307X and Y416X showed higher expression compared to wild-type (WT) FOXG1. For missense variants, I194S, F215L, R230H, and G252V displayed lower expression, while others (P182Q, N187K, L235H, and G252D) exhibited levels similar to or higher than WT FOXG1. All frameshift variants (Q86Dfs*34, Q86Rfs*106, W255Lfs*156) consistently displayed very low expression (Fig. 1B–D).
Protein expression levels were categorized as very low (Q86Dfs*34, Q86Rfs*106, E136X, R203H, G252V, W255Lfs*156), low (I194S, F215L), normal (P182Q, N187K, L235H), and high (G252D, Y416X, Y307X). Interestingly, variants with very low expression correlated with moderate-to-severe MRI findings, suggesting a link between reduced expression and severe phenotypes. This assay alone identified 54.5% (6/11) of cases with moderate-to-severe brain anomalies. However, due to its low correlations with the brain MRI and clinical severity scores (R2 = 0.0072 and 0.046, respectively; Supplementary Fig. 3A), additional assays are necessary for a more accurate prediction of FOXG1 syndrome severity.
Failure to repress the COUP-TFI promoter is associated with moderate-to-severe brain anomaliesGiven FOXG1’s established role in repressing COUP-TFI (NR2F1) expression [7], we analyzed Foxg1 and Nr2f1 expression using our scRNA-seq data from mouse cortices at E13.5 and E15.5 [30]. Additionally, scRNA-seq data from mouse embryos spanning E10–E18 were examined [31] (Fig. 2, Supplementary Fig. 4). Cells were categorized into three main types: progenitor (i.e., radial glial cells), precursor (i.e., intermediate progenitors), and neuron, based on various cell markers, including Pax6, Emx1, Eomes, Neurod6, Tbr1, Bcl11b, Satb2, and Fabp7 (Fig. 2A) [30]. We found that, at both stages, Foxg1 was predominantly expressed in neurons, with minor expression in progenitors and precursors, while Nr2f1 showed neuron-specific expression (Fig. 2B). Interestingly, Foxg1 and Nr2f1 expression appeared mutually exclusive (Fig. 2C), consistent with Foxg1’s role in repressing Nr2f1 expression during cortical development [7]. We extended this analysis using scRNA-seq data from the human embryonic forebrain at GW18 [32], where we observed a similar mutual exclusion between FOXG1 and NR2F1, confirming the conserved nature of this regulatory mechanism between mouse and human cortical development.
Fig. 2: Expression of Foxg1 and COUP-TFI (Nr2f1) in the developing cerebral cortex and the COUP-TFI repression ability of FOXG1 variants.
A UMAP plots depicting scRNA-seq results of the cortices from mouse embryos at E13.5 and E15.5 (top); mouse embryos at E10, 12, 14, 15, 16 and 18 (middle); and human fetus at GW18 (bottom). Three developmental states (progenitors, precursors, and neurons) are identified based on their marker genes. B Heat maps illustrating the expression of Foxg1/FOXG1 (red) and COUP-TF1 (Nr2f1/NR2F1, green), revealing distinct expression patterns. Foxg1/FOXG1 and Nr2f1/NR2F1 expression exhibit mutually exclusive expression patterns. C Scatter plots of Foxg1/FOXG1 and Nr2f1/NR2F1 expression in these cells shows relatively few cells co-expressing Foxg1/FOXG1 and Nr2f1/NR2F1. D Diagram outlining the luciferase reporter assay. Luciferase expression is driven by the COUP-TFI promoter (COUPTF1pro) and its enhancer-like region PBS1. Co-transfection of Foxg1 and the reporter construct allows FOXG1 to bind to the PBS1 region, repressing the expression of downstream genes. E Verification of WT FOXG1 repression of COUP-TFI, requiring PBS1. Cells transfected with Foxg1 along with the Luciferase reporter driven by COUPTF1pro without PBS1 exhibited high luciferase activity. In contrast, the inclusion of PBS1 reduced the luciferase activity, confirming a repression function of FOXG1 (n = 3 independent transfections). F Luciferase activity of cells transfected with Foxg1 variants along with the luciferase reporter driven by the COUP-TFI promoter and PBS1. While the expression of Y416X, L235H, Y307X, P182Q, G252D, and F215L resulted in similarly low luciferase activity as WT FOXG1, the expression of other variants showed significantly higher luciferase activity, indicating impaired COUP-TFI repression function (n = 3 independent transfections). Error bars: S.E.M. *p < 0.05, **p < 0.01, ***p < 0.001. ****p < 0.0001. One-way ANOVA; post-hoc: Uncorrected Fisher’s LSD.
To assess the impact of FOXG1 variants on the ability to repress COUP-TFI expression, we used a luciferase reporter construct with the COUP-TFI promoter and COUP-TFI enhancer PBS1 (Fig. 2D). Previous studies showed FOXG1 binding to PBS1 represses the promoter activity, downregulating COUP-TFI [7]. In this “COUP-TFI repression assay,” the reporter construct and Foxg1 cDNA were co-transfected into U87-MG cells. WT FOXG1 led to low luciferase activity, consistent with its repressor role in COUP-TFI expression (Fig. 2E) [7].
Comparing luciferase luminescence among cells expressing FOXG1 variants, 4 variants (P182Q, L235H, Y307X, and Y416X) showed low activities similar to WT FOXG1 (Fig. 2F). In contrast, G252D and F215L, associated with mild MRI severity, exhibited slightly increased activity, suggesting partial LOF. The remaining 8 FOXG1 variants, associated with moderate (E136X, N187K, I194S, R230H, G252V) or severe (Q86Dfs*34, Q86Rfs*106, W255Lfs*156) MRI severity scores, displayed elevated activity, indicating LOF in COUP-TFI repression. Notably, all 6 variants with very low protein expression exhibited significantly higher luciferase activity compared to WT FOXG1 (p < 0.05, Fig. 2F), consistent with expected LOF effects. Among other variants, only N187K and I194S showed LOF in the repression of luciferase activity, both associated with moderate MRI severity, regardless of their expression levels (N187K: normal; I194S: low). While correlations between COUP-TFI repression and the brain MRI (R2 = 0.085) and clinical severity (R2 = 0.090) scores remained low (Supplementary Fig. 3B), this additional assay increased sensitivity in detecting variants causing moderate-to-severe brain anomaly from 54.5% (6/11) to 72.7% (8/11).
Impact on neuronal migration correlates with brain MRI severityPreviously, FOXG1 overexpression in the developing mouse cortex at E13.75 delayed neuronal migration and attenuated layer IV neuronal fate [7]. To investigate how FOXG1 variants impact these functions, we used IUE to introduce constructs overexpressing WT FOXG1 or its variants, along with GFP plasmid, into neural progenitors at E13.75 (Fig. 3). Three days post-IUE, brain sections stained with TBR1 (T-Box Brain Transcription Factor 1) as a marker for the deep cortical plate (CP) showed that, in control brains electroporated with the empty vector, about half of the GFP+ cells had migrated from the ventricular zone (VZ) to the CP, with some cells in transit within the intermediate zone (IZ) and VZ. Notably, upon electroporation of WT Foxg1, most electroporated GFP+ cells were distributed in the IZ and VZ, with very few cells (<10%) reaching the CP, consistent with previous studies [7].
Fig. 3: Effects of overexpression of FOXG1 WT and variants on neuronal migration by IUE.
A Neuronal distribution of brains electroporated with Foxg1 and the pathogenic variants. Mouse brains were electroporated with GFP along with Foxg1 WT or its variants at E13.75. Neuronal cell distribution was examined 3 days after IUE in brain slices stained with the layer marker TBR1 (red) and DAPI (blue). While more than half of the cells electroporated with the empty vector migrated to the CP, FOXG1 overexpression altered neuronal migration, causing most cells to accumulate in the VZ and IZ. Cells electroporated with pathogenic Foxg1 variants displayed varying degrees of migration alteration. Bar = 100 μm. B The bar graph shows cell distributions in the VZ, IZ, and CP after IUE. C The bar graph shows cell distributions in the CP after IUE. Dashed horizontal lines indicate 10 and 25% of cells distributed in the CP. Error bars: S.E.M. *p < 0.05, **p < 0.01, ***p < 0.001. ****p < 0.0001. Two-way ANOVA, post-hoc: Dunnett’s multiple comparisons test (n = 3 independent IUE experiments).
IUE of disease-associated FOXG1 variants showed varied effects (Fig. 3). Brains electroporated with 6 variants with very low expression (Q86Dfs*34, Q86Rfs*106, E136X, R203H, G252V, W255Lfs*156) showed many GFP+ cells in the CP (>25%), similar to brains electroporated with the vector only, indicating functional impairment. Among the two other variants showing LOF in COUP-TFI repression, I194S overexpression also resulted in > 25% of cells reaching the CP. Curiously, N187K overexpression led to very few cells reaching the CP (<10%), suggesting retained functionality in affecting neuronal migration. Of the remaining 6 variants without LOF in COUP-TFI repression, P182Q and Y307X exhibited similar percentages of cells in the CP compared to WT Foxg1 (<10% in all 3 groups, p > 0.05), suggesting relatively normal function. F215L and G252D resulted in 10–25% of cells migrating to the CP, suggesting some LOF. L235H and Y416X led to > 25% of GFP+ cells in the CP, suggesting a greater LOF. Collectively, our findings reveal a good correlation between functional impairment in these variants and the severity of brain anomalies (R2 = 0.405, Supplementary Fig. 3C). Combining these three functional assays increased the prediction rate for variants causing moderate-to-severe brain anomaly to 90.9% (10/11), except for the Y307X variant, which, despite minimal LOF in all three assays, was associated with severe brain anomalies.
Correlation between brain MRI severity and altered cell fateTo investigate the impact of FOXG1 variants on neuronal cell fate, brains after IUE at E13.75 were stained with layer-specific markers at P7, including BRN2 (Brain-specific homeobox/POU domain protein 2) for layer II/III and RORβ (Retinoic acid-related orphan receptor beta) for layer IV (Fig. 4). In control brains, most GFP+ cells reached layer IV and expressed RORβ but not BRN2. In contrast, FOXG1 overexpression shifted cells to layers II/III, V, and VI, predominantly RORβ- and BRN2+, consistent with a mutually repressive interaction between RORβ and BRN2 [41].
Fig. 4: Effects of the expression of FOXG1 and its variants on the expression of the layer II/III and IV neuronal markers in the developing cortex.
Mouse brains were electroporated with GFP (green) along with Foxg1 variants or the empty vector at E13.75. Brain sections at P7 were immunostained with the layer II/III marker BRN2 or layer IV marker RORβ, and counterstained with DAPI (blue). A Neurons electroporated with the empty vector were mostly BRN2- and distributed in layer IV, while FOXG1-expressing neurons were redistributed to layer II/III and expressed BRN2 (red). Neurons expressing pathogenic Foxg1 variants displayed variable effects on this fate change. The lower panels show high-magnification images with separated channels. Arrows indicate BRN2+ neurons. Bars = 100 μm. The bar graph shows the percentage of BRN2+ neurons among GFP+ cells after overexpressing Foxg1 WT and its variants. The percentage of BRN2+ cells inversely correlates with the severity of brain malformations. B Conversely, most neurons electroporated with the empty vector were RORβ+ (red), while neurons expressing FOXG1 were redistributed to layer II/III and became RORβ-. Neurons expressing pathogenic variants displayed varying effects on this fate change. Lower panels show the high-magnification images with separated channels. Arrows indicate RORβ+ neurons. Bars = 100 μm. The bar graph shows the percentage of RORβ+ among GFP+ cells after overexpressing FOXG1 WT and its variants. The percentage of RORβ+ cells correlates well with the severity of brain malformations. Error bars: S.E.M. *p < 0.05, **p < 0.01, ***p < 0.001. ****p < 0.0001. One-way ANOVA, post-hoc: Uncorrected Fisher’s LSD (n = 3 independent IUE).
Brains electroporated with Foxg1 variants causing normal (P182Q), mild (F215L), and moderate (N187K) brain anomalies showed a slight but significant reduction in BRN2+ cells compared to Foxg1 WT-electroporated brains (Fig. 4), indicating some LOF in altering neuronal differentiation. Notably, brains electroporated with variants leading to severe brain anomalies (L235H and W255Lfs*156) exhibited even greater reductions in BRN2+ cells, suggesting disrupted FOXG1 function. Conversely, brains electroporated with normal-to-moderate variants (P182Q, F215L, and N187K) showed partial LOF in directing neurons from RORβ+ to RORβ-, whereas severe variants (L235H and W255Lfs*156) lost most of this function (Fig. 4). Although we assessed only a subset of the 14 variants spanning different severity levels due to the complexity of this assay, these results suggest a correlation between the ability to alter cell fate and the severity of brain anomalies in FOXG1 syndrome.
Clinical severity prediction workflow for FOXG1 syndromeBased on these functional assays, we developed a flowchart to predict the severity of brain anomalies resulting from FOXG1 variants (Fig. 5A). FOXG1 variants initially classified as very low expressers (<50% of WT expression) typically lead to severe (Q86Dsf*34, Q86Rfs*106, and W255Lfs*156) or moderate (E136X, R230H, and G252V) brain anomalies. This assay identified 54.5% (6/11) of moderate-to-severe cases and no (0/2) mild cases. Further refinement is achieved through a COUP-TFI repression assay. For variants with normal or high expression levels, those exhibiting impaired COUP-TFI repression ability (N187K and I194S) resulted in moderate brain anomalies, increasing the sensitivity of detecting moderate-to-severe cases from 54.5% (6/11) to 72.7 (8/11). FOXG1 variants demonstrating relatively normal COUP-TFI repression ability should undergo a further assessment of their impact on neuronal migration through IUE. In cases of WT and variants leading to normal MRI (P182Q), fewer than 10% of cells typically reach the CP. Variants leading to severe (L235H) and moderate (Y416X) brain anomalies usually allow a higher percentage of cells (>25%) to reach the CP, while mild variants (e.g., F215L and G252D) result in 10–25% of cells reaching the CP. Among these variants, only Y307X, associated with severe brain anomalies, does not align with our prediction model. Combining all three assays resulted in a sensitivity of 90.9% (10/11) for moderate-to-severe cases, 100% (2/2) for mild cases, and an overall sensitivity of 92.3% (12/13).
Fig. 5: A flowchart for predicting the severity of brain anomalies caused by FOXG1 variants through in vitro and in vivo assays.
A The expression levels of FOXG1 variants are first examined, and variants with very low expression (<50%) tend to cause severe to moderate brain anomalies. This categorization can be further refined through the COUP-TFI repression assay. While all very low expression variants exhibit impaired repression, among variants with normal to high expression levels, those showing impaired COUP-TFI repression tend to lead to moderate brain anomalies. For FOXG1 variants demonstrating relatively normal COUP-TFI repression ability, their effects on neuronal migration and differentiation are further assessed through IUE. In cases of WT FOXG1 and variants that are associated with normal MRI, fewer than 10% of cells typically reach the CP. Variants leading to severe and moderate brain anomalies usually result in a higher percentage of cells (>25%) reaching the CP, while mild variants lead to 10–25% of cells reaching the CP. It’s worth noting that among these variants, only Y307X, which is associated with severe brain anomalies, does not align with our prediction model. #: variants that did not entirely fit the model. B PCA of FOXG1 variant severity. Data from protein expression, COUP-TFI repression, and neuronal migration assays were plotted in a three-dimensional scatter plot, with severity groups color-coded. Clustering analysis effectively differentiates normal, mild, moderate, and severe brain anomalies, with the exception of N187K and Y307X, which remain outliers.
Interestingly, there are only moderate correlations between these 3 assays (Supplementary Fig. 5), suggesting each functional assay provides valuable insights into the properties and functions of different variants. To further validate our workflow, we applied principal component analysis (PCA) to visualize the clustering of variants based on FOXG1 protein expression, COUP-TFI repression in the luciferase assay, and the percentage of cells reaching the CP in the migration assay. This analysis revealed a clear separation of normal, mild, moderate, and severe MRI phenotypes, with the exception of N187K and Y307X (Fig. 5B), reinforcing the robustness of our predictive framework.
Comments (0)