
Remember me

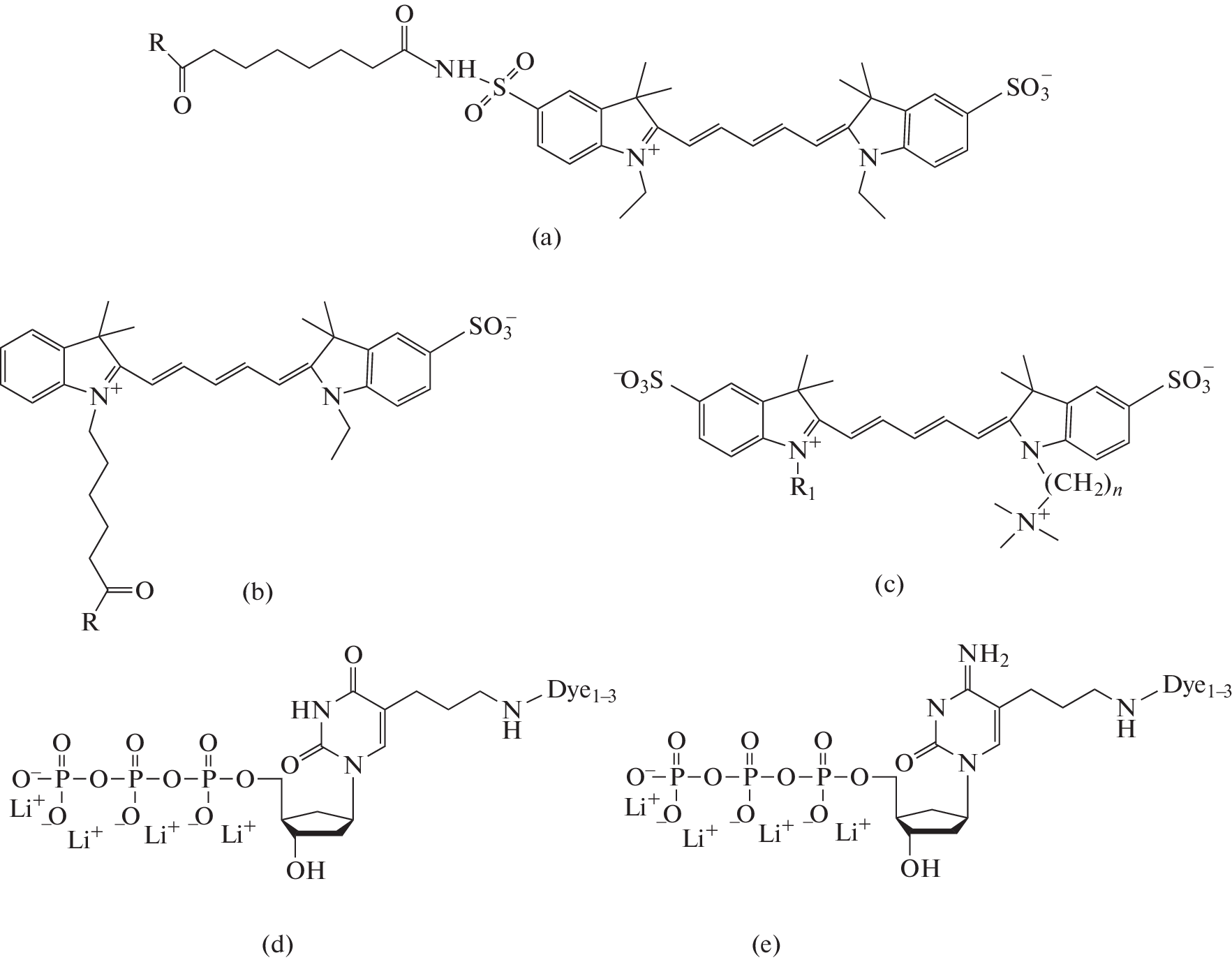
In this work, the substrate behavior, namely, inhibition and efficiency of incorporation into the growing DNA chain during PCR of six different pairs of zwitterionic electroneutral deoxynucleoside triphosphates (dU and dC) with similar modifications on two DNA templates with significantly different GC compositions, was studied. The reaction was carried out with incomplete replacement of natural triphosphates with fluorescently labeled analogues in accordance with the previously found average optimum concentration (varies for different derivatives) [8]. Modifications introduced into heterocyclic bases differed in the length of the linkers and the structure of the fluorophore. Two control pairs were also taken for comparison, differing in the structure of their Cy5 fluorophores from the four pairs studied. These pairs are laboratory standards, as they are used as the main reagents for introducing a label into DNA in the biological microarray technology [9].
This work used the real-time PCR method with EvaGreen staining to exclude signals from the introduced fluorescent Cy5 labels. The averaged fluorescence signal accumulation curves from both templates appear as follows (Fig. 2). The amplification efficiency was determined from the obtained amplification signal accumulation curves E [12, 13], given in Table 2.
Fig. 2.
Rate of accumulation of the PCR product on the (a) AT-rich template of S. aureus and (b) GC-rich template of M. tuberculosis on the example of dUss and dCss. The control is the sample without modified deoxyribonucleoside triphosphates.
Table 2. Amplification efficiency E*, embedding coefficient Ki**, product yield, and incorporation density on templates of S. aureus and M. tuberculosis and six pairs of deoxyribonucleoside triphosphate derivativesThe signal accumulation curves during PCR show that on both templates the amplification efficiency values for the cytidine analogues exceed the values for the uridine analogues. On the AT-rich template, a significantly greater difference in the signal accumulation rate can be seen.
The amplification products were purified using microcolumns to remove fluorescently labeled nucleotides that were not incorporated into the DNA chain from the mixture and to avoid their influence on the total absorption during subsequent spectrophotometric measurements to calculate the incorporation efficiency. Using electrophoretic analysis, purified PCR products were checked for the absence of amplification by-products and the length of the PCR product was compared with its theoretical (calculated) length. The efficiency of label incorporation in the resulting purified product was then determined spectrophotometrically by measuring the absorption of the PCR product solution at wavelengths of 260 and 647 nm (for the total absorption of DNA and fluorophore, respectively), and the values of the incorporation coefficient, incorporation density, and specific incorporation density were calculated. Amplification followed by purification of the product and the optical density measurements were performed in triplicate for each fluorescently labeled deoxynucleoside triphosphate.
Embedding coefficient Ki was introduced as a rapid method for assessing the substrate efficiency of fluorescently labeled deoxynucleoside triphosphates and was calculated as a simple ratio of the optical density at 647 nm to the optical density at 260 nm for each sample. The incorporation density was calculated as the ratio of the molar amount of label to the molar yield of DNA product. The specific incorporation density was calculated as the ratio of the molar amount of label in the sample to the molar amount of the same nucleotides in the resulting PCR product.
The average values obtained during the experiments, calculated from three replicates, are presented in Table 2.
From Table 2 it is evident that on both templates in almost all cases the amplification efficiency (E) in the presence of modified cytidine is greater than in the presence of uridine. But at the same time, it is noticeable that the difference in the obtained values is small within each pair; on this basis, it can be concluded that the efficiency of amplification depends weakly on the nature of the fluorescently labeled nitrogenous base (dU and dC) introduced into the reaction. This may be due to incomplete substitution of natural triphosphates in the reaction mixture (16 μM labeled and 200 μM natural).
Embedding coefficients Ki, in contrast to amplification efficiency E, differ more; they are larger for dU. One can see a great difference when comparing different bases in one pair. On a GC-rich template, fluorescently labeled cytidines have a greater opportunity to be incorporated into the chain due to the nucleotide composition of the template, i.e., a greater number of potential “landing sites.” However, the high efficiency of incorporation of deoxyuridines leads to the fact that on the GC-rich template the values of the incorporation coefficients become close for dU and dC. It is important to note that Ki should be used to compare the efficiency of incorporation of identical nucleotides, since these coefficients depend on the GC composition of the template.
However, for a quick assessment of the efficiency of incorporation on “polar” GC-composition templates, a comparison of the dU/dC ratio (in the case of an AT-rich template) and dC/dU (for a GC-rich template) is very useful. It is evident that on the AT-rich template the efficiency of incorporation of uridine analogues is much higher than that of cytidine analogues, while on the GC-rich template the superiority of the values of the ratio of incorporation coefficients dC/dU is leveled by the higher efficiency of incorporation of uridines.
The specific incorporation density index is suitable for comparing the substrate properties of modified dU and dC, since it takes into account the number of identical nucleotides in the target DNA. This indicator also suggests that fluorescently labeled uridine derivatives are better integrated into the DNA chain, since in most pairs their values are higher than those of cytidine analogues.
When analyzing the incorporation coefficient, one can see the influence of the structure of triphosphates and the composition of the template. Higher values were obtained on the template of S. aureus for control samples dUc1 and dUc2, which we currently use as primary labels when conducting microarray analysis. It can be noted that, in the four pairs studied, the best incorporation into the chain is observed on both templates for samples with a long linker between the fluorophore and the nitrogenous base.
In the case of the AT-rich template, the values of the incorporation coefficient in pairs are significantly higher than those of the uridine analogues. On the GC-rich template, the advantage of embedded cytidines is visible in terms of the efficiency values, and also, in most cases, in terms of the values of the incorporation coefficients, but the difference in values is small.
Based on the data obtained, it can be concluded that uridine derivatives are the best substrates for templates of different GC composition due to their significantly higher efficiency of incorporation compared to cytidines. The length of the linker between the fluorophore and the nitrogenous base increases the efficiency of incorporation of both deoxynucleosides. A higher incorporation density is accompanied by greater inhibition of the reaction, which, apparently, indicates the involvement of the modified substrate in a complex with the polymerase with some complications and, as a consequence, a slowdown in the kinetics of amplification. Conversely, less involvement in the reaction as a substrate results in less inhibition of the reaction by such derivatives.
Comments (0)