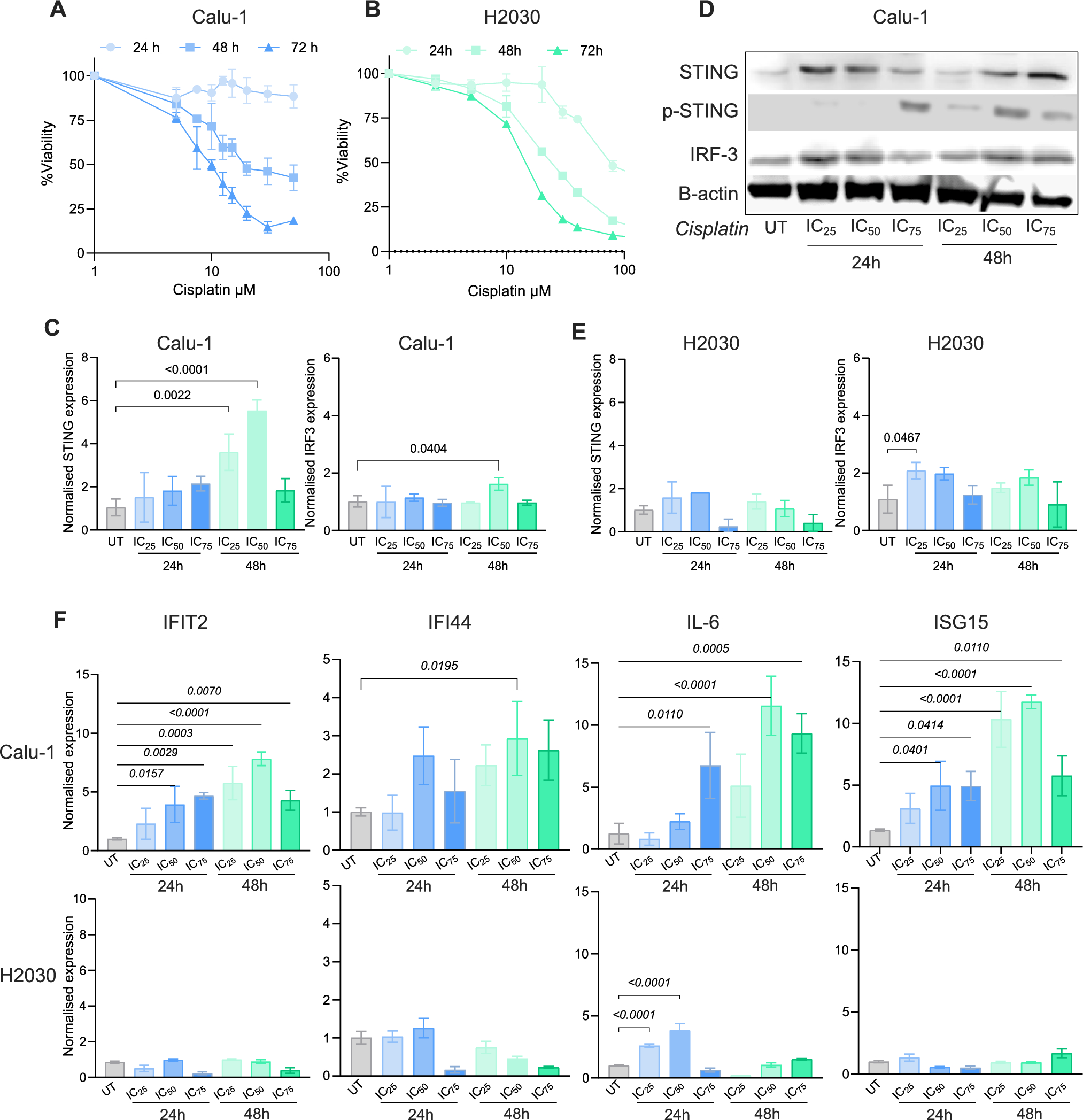
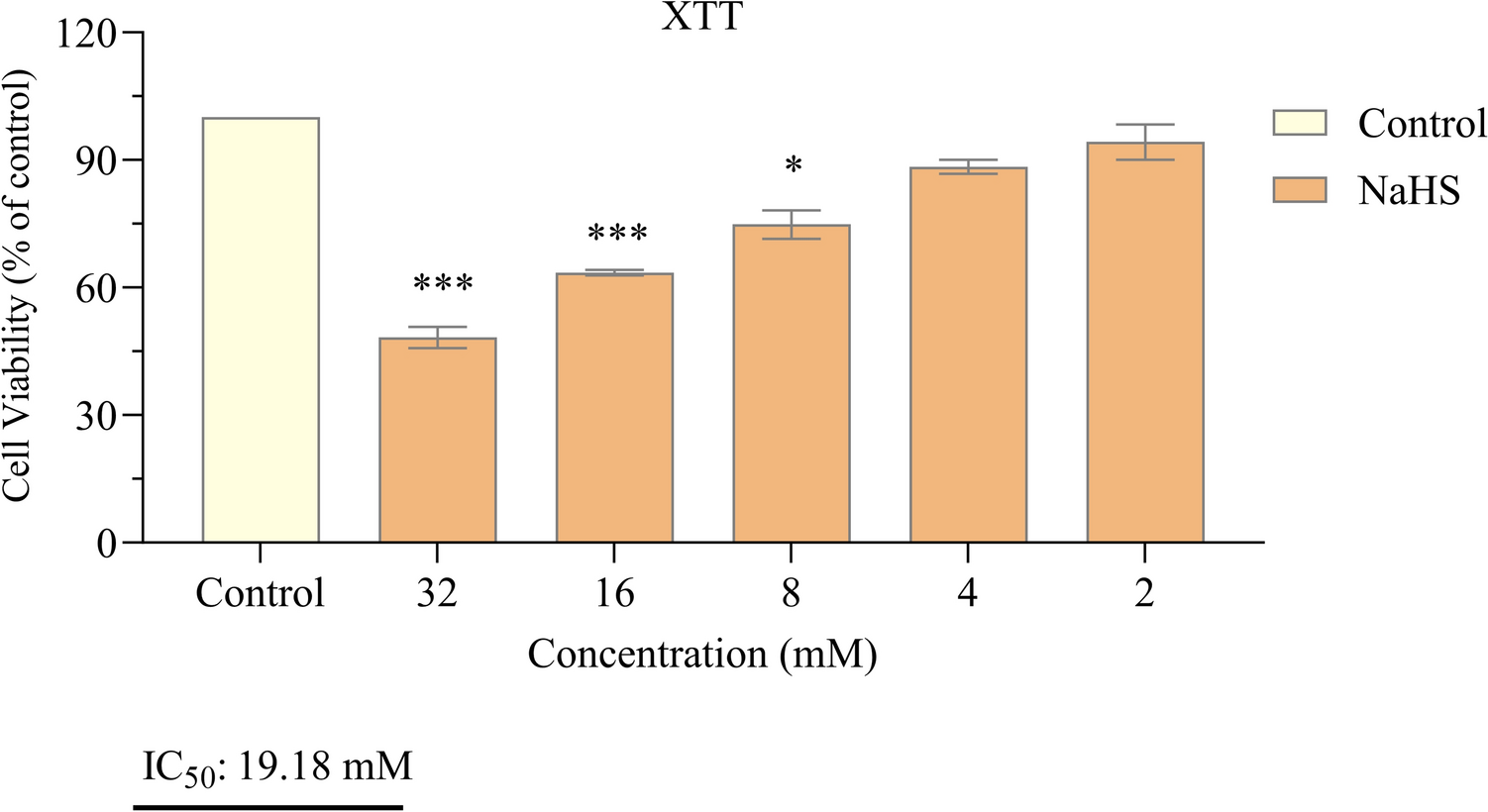
The relationship between NaHS and H₂S in cancer is complex; the effects of H₂S can both inhibit cell growth and enhance cell proliferation. This bidirectional effect is still under investigation, and no definitive conclusions have been reached regarding the role of H₂S in cancer therapy. Therefore, additional research is required to investigate the potential application of NaHS and hydrogen sulfide in cancer therapy. In this study, we investigated the effects of NaHS used as H₂S donor on cytotoxicity, growth signaling pathways, and programmed cell death (apoptosis) in the SH-SY5Y cell line, a derivative of neuroblastoma, in vitro. The experimental data obtained revealed that NaHS significantly inhibited the proliferation of SH-SY5Y cells in a concentration-dependent manner. The IC50 value was determined to be 19.18 mM following 24 hours of exposure to NaHS. In their study investigating the anticancer activity of NaHS, Zhang et al. found that cell viability in SGC7901 gastric cancer cells decreased in a concentration-dependent manner, which is consistent with our findings [22]. In contrast, Liu et al. reported that the exogenous hydrogen sulfide donor NaHS could induce the proliferation and invasion of cells by increasing the expression levels of MMP-2 and MMP-9 in human bladder cancer EJ cells [23]. Research on colorectal cancer cells have demonstrated that exogenous H2S (NaHS) promotes cell proliferation at a concentration of 200 μM in the HCT116 cell line, while concurrently decreasing cell survival at concentrations between 50 and 200 μM in the WiDr cell line [22, 23]. These findings suggest that the effects of different dosages and methods of H2S exposure are likely specific to the cell line under investigation. Based on these findings, a bell curve model was proposed to explain the bidirectional effects of H₂S on cancer cells. According to this model, endogenous H₂S or low levels of exogenous H₂S may have a protective effect against cancer cells, while high concentrations or extended exposure to H₂S may result in the death of cancer cells [24]. Lee et al. conducted a study examining the effects of exogenous H₂S donors, specifically NaHS and GYY4137, on both cancerous cell lines (MCF7 and HepG2) and non-cancerous cell lines (MCF10A and WI38). The study revealed that long-term exposure to low concentrations of exogenous H2S exhibited selective anticancer effects when compared to a single exposure. Mechanistic analyses have demonstrated that H2S disrupts pH regulation by increasing the production of metabolic acids in cancer cells, ultimately leading to cell death. These findings indicate that H2S holds significant potential as a selective anticancer agent [25]. In a study, GYY4137, H2S donor, was found to inhibit tumor growth by arresting the cell cycle and promoting apoptosis. GYY4137 demonstrated its ability to suppress tumor growth in hepatocellular carcinoma (HCC) cells and in a HepG2 xenograft model by targeting the STAT3 pathway. This effect is mediated through the reduction of phosphorylated STAT3 (p-STAT3) levels, as well as the inhibition of the cell cycle and angiogenesis [26]. However, some studies indicate that NaHS exhibits cytoprotective rather than cytotoxic effects on certain non-malignant cell types under specific conditions. These findings reinforce NAHS's biosafety profile. For example, Denizaltı et al. reported that NaHS enhances cell viability, proliferation, and migration in L929 mouse fibroblast cells. Additionally, it has been demonstrated that NaHS promotes wound healing by reducing oxidative stress under hyperglycemic conditions and may serve as a potential therapeutic agent for skin repair [27]. Similarly, Yu et al. reported that NaHS exhibited protective effects against hemin-induced ferroptosis in BV2 microglial cells. The effect was linked to CBS/H₂S-mediated oxidative stress reduction and increased GPX4, SLC7A11, and GSH levels [28]. These findings suggest that NaHS may not exert cytotoxic effects on certain non-malignant cell types. Supporting this, several studies have demonstrated the cytoprotective and antioxidant effects of NaHS specifically in SH-SY5Y neuroblastoma cells. Whiteman et al. (2005) reported that NaHS (25–125 µM) significantly reduced HOCl-mediated cytotoxicity and oxidative damage in SH-SY5Y cells [29]. Likewise, Whiteman et al. (2004) demonstrated that NaHS (50–250 µM) prevented peroxynitrite-induced protein nitration and cytotoxicity in the same cell line [30]. Koike et al. (2017) further showed that NaHS (200 µM) protected SH-SY5Y cells from methylglyoxal-induced toxicity by activating Keap1/Nrf2 pathways and increasing intracellular glutathione levels [31]. Additionally, Tiong et al. (2010) indicated that NaHS (100–1000 µM) exerted protective effects against 6-hydroxydopamine-induced cell damage through PKCa and PI3K/Akt pathway activation in SH-SY5Y cells [32]. In line with both the literature and our findings, NaHS exhibited protective effects on SH-SY5Y cells at low concentrations, whereas it exerted antiproliferative effects at higher concentrations. A limitation of this study is that the effects of NaHS on healthy cells were not directly investigated. However, previous studies have reported that NaHS may have protective and antioxidant effects in non-malignant cell types. Future studies should include healthy cells to better evaluate the safety and therapeutic potential of NaHS.
To investigate the effect of NaHS on the biological responses of SH-SY5Y cells, we analyzed the expression levels of various proteins involved in growth signaling and apoptosis. EGFR, a member of the receptor tyrosine kinase family, plays a crucial role in various cellular processes and is regarded as a key target for cancer treatment. The activation of EGFR is linked to the proliferation, migration, and invasion of neoplastic cells, and it is found to be overexpressed in several cancer types, including SH-SY5Y neuroblastoma cells [5, 10]. Phosphorylation of the EGFR can activate several signaling pathways, including the ERK1/ERK2, PI3K/AKT/mTOR, and MAPK/MEK/ERK pathways [25, 26]. In a recent investigation, it was observed that NaHS facilitated the phosphorylation of AMP-activated protein kinase (AMPK), inhibited the mTOR signaling pathway, and promoted autophagy in colon epithelial cells [33]. Wu et al. observed that concentrations of 25–50 μM NaHS increased the phosphorylation of EGFR and ERK1/2, as well as angiogenesis, in hepatocarcinoma cell lines (SMMC-7721 and Huh-7), but these phosphorylation and angiogenesis diminished at concentrations of 800–1000 μM NaHS. H₂S had no significant effect on growth, migration, apoptosis, or protein expression levels related to the PTEN/AKT and EGFR/ERK/MMP-2 signaling pathways in normal human liver cells (L02). These findings suggest that H₂S may modulate the migration and growth of hepatocellular carcinoma cells through the EGFR/ERK/MMP-2 signaling pathway [34]. Similar to our findings, NaHS inhibited the EGFR, HER2, ERK1, and ERK2 signaling pathways in the SH-SY5Y cell line. These results indicate that NaHS influences cellular functions in neuroblastoma cells by modulating signaling pathways and that this mechanism contributes to its anticancer activity. mTOR is a kinase that regulates cellular growth, metabolism, and autophagy. Abnormal activation of mTOR is particularly associated with uncontrolled cell proliferation and resistance to apoptosis in cancer [35]. Therefore, mTOR represents a significant therapeutic target for various cancers, including neuroblastoma. However, in our study, we observed that NaHS did not alter mTOR levels. This does not imply that mTOR is not involved in the antiproliferative and pro-apoptotic effects of NaHS. While NaHS decreased upstream signaling molecules such as HER-2, EGFR, and ERK, it may not affect the total level of mTOR. This suggests that NaHS may exert its effects through upstream pathways rather than through direct modulation of mTOR. In addition, the potential changes in the levels of phosphorylated mTOR (p-mTOR), the active form of mTOR, should be considered. Indeed, studies conducted by Wang et al. (2020) and Yu et al. (2009) have revealed that the biological effects and clinicopathological associations of total mTOR and p-mTOR may differ [33, 34]. Therefore, more comprehensive studies are necessary to fully elucidate the role of the mTOR signaling pathway in this process.
Apoptosis is the intrinsic cell death program and serves as a crucial mechanism for maintaining normal developmental processes and tissue homeostasis in multicellular organisms [20, 36]. Apoptosis and the expression levels of caspase-3, a crucial protein associated with apoptosis, were analyzed in detail. Our results indicated that 19.18 mM NaHS significantly enhanced apoptosis. HER2 signaling is primarily mediated by PI3K [37]. The PI3K/AKT pathway plays a crucial role in the metabolism, migration, proliferation, and apoptosis of cancer cells [38, 39]. Wu et al. reported that the BAX/BCL-2 ratio increased when this signaling pathway was inhibited using high concentrations of NaHS [34]. Similarly, Murata et al. investigated the induction of apoptosis in the oral cancer cell line Ca9-22 by H2S. H2S significantly induced apoptosis in Ca9-22 oral cancer cells. In Ca9-22 cells, H2S was found to enhance the expression of a protein known as PHLDA1. This protein subsequently activates caspase-3 during apoptosis, ultimately resulting in cell death [40]. In accordance with previous studies, our findings, based on the data obtained by flow cytometry and ELISA methods, demonstrate that the exogenous H2S donor NaHS induces apoptosis in SH-SY5Y neuroblastoma cells. One of the limitations of this study is the absence of caspase-8 and caspase-9 activity measurements, which are crucial for distinguishing between the intrinsic and extrinsic apoptotic pathways. Future research will aim to address this gap.
In conclusion, the suppression of growth signaling pathways by NaHS resulted in a significant increase in caspase-3 levels. These findings suggest that NaHS may operate through a mechanism that enhances apoptosis by inhibiting cell growth in SH-SY5Y cells. This modulation of growth signaling pathways offers a deeper understanding of the effects of NaHS on cancer cells. This study is limited to in vitro cell models, and in vivo validation is necessary. Additionally, further research is needed to investigate the long-term effects and potential toxicities of NaHS. As a next step, we plan to validate our findings in in vivo models and investigate additional apoptotic and proliferative markers. We also aim to explore the potential of NaHS in combination therapies with standard treatments, considering the complex resistance mechanisms in neuroblastoma. Additionally, further research is needed to investigate the long-term effects and potential toxicities of NaHS.




Comments (0)