
Remember me

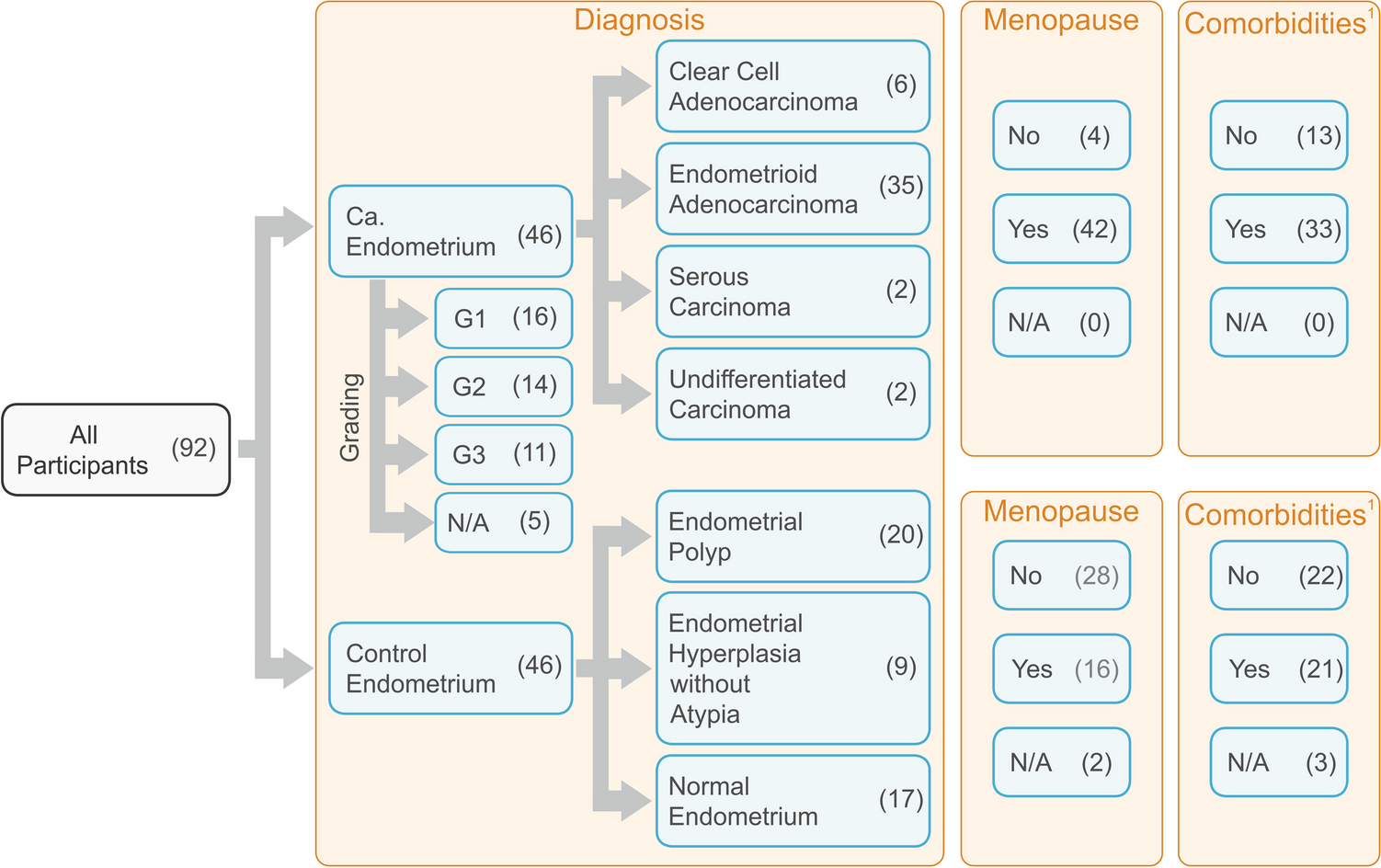
EC represents one of the most frequent malignancies in females, and in 2020, it was the 14th most common cause of cancer-related mortality worldwide. An umbrella review conducted in 2018 revealed that obesity is the major risk factor for EC in both pre-menopausal and post-menopausal women. However, nulliparity, diabetes, and other hormonal and metabolic pathways involved in adiposity may also contribute to EC development (Raglan et al. 2019).
Moreover, recent studies emphasize that the effectiveness of EC therapy relies on the cellular and molecular mechanisms that interplay with the survival and resistance of tumor cells. During the tumor growth, the oxygen concentration and nutrient accessibility are decreased, which leads to hypoxia. Next, it activates angiogenesis-related and hypoxia-inducible factors, which are involved in the regulation of a vast number of genes. As a result, tumor cells must adapt to unfavorable conditions that may promote cancer metastasis and invasiveness (Salinas-Vera et al. 2022). Thus, we analyzed the HIF1A, EPAS1, and VEGFA gene expression patterns in cancerous and normal endometrial tissues involved in hypoxia, angiogenesis, and other biological processes and ultimately play a role in EC.
It is a dogma that numerous genes are involved in endometrial malignancies. The multiplicity of genes could illustrate the complexity of this issue, proteins, and other factors involved in the EPAS1, HIF1A, and VEGFA-related pathways (Supplementary Materials, Figure S1). It was shown that, e.g., EPAS1 at the regulomics level has at least 31 upstream regulators and directly interacts with 27 proteins. Its expression is associated with more than ten cancer types, and its regulation on protein level is complex in physiological conditions and especially in malignancies (Kristan et al. 2021). The HIF1A regulation pathway is even more complex. Over 85 miRNAs have been reported to target this gene, and HIF1A downstream targets include protein-coding genes, long non-coding RNAs, and miRNAs (Kunej 2021).
In clinical practice, it would be almost impossible to search for diagnostic or prognostic candidate genes in the therapy of the future, so it is worth looking for only a few target genes. Thus, this work focused on angiogenesis, the hallmark of cancer growth, hypoxia, an angiogenesis driver, and three related to those processes genes (Fig. 13).
Fig. 13
Map of the HIF1A, EPAS1, and VEGFA interplay (cpb.molgen.de)
Case–control studyThe case–control study showed significant differences in age, BMI, and the number of comorbidities. These results, although not surprising, indicate that the group we studied is similar to the subpopulations analyzed by other authors (Nevadunsky et al. 2014). Moreover, as described by other authors, older women with EC were associated with more aggressive disease features, limited surgical or adjuvant therapy treatment, and a worse prognosis. Age could be a dependent or independent predictor of EC outcome. It was established that the higher age is related to a higher risk of disease recurrence and EC-specific mortality (Hag-Yahia et al. 2021; Biomorfologi et al. 2022). Age is not a modifiable factor, but aging affects the expression of genes. However, in our research, we did not study this issue.
Additionally, it was confirmed that obesity was associated with earlier EC occurrence or time of diagnosis (Nevadunsky et al. 2014; Gao et al. 2016; Park 2022). Metabolic and endocrine effects of obesity on EC susceptibility could be the explanation. Both higher BMI and age at diagnosis were observed to be strongly associated with EC (Nevadunsky et al. 2014). Primarily, obesity was confirmed to be positively related to the incidence of developing EC, particularly in perimenopausal women (Gao et al. 2016), but negatively in the early stage regardless of cancer subtypes and menopausal status (Gao et al. 2018). It is especially hazardous as patients are usually unaware of the obesity-related risk factor and BMI-related risk-reducing strategies. Women who lose weight have a better response to therapy and may also have improved oncological outcomes and result in fewer diagnosed comorbidities (Barr et al. 2021). Thus preventive for EC weight control should be taken as a standard approach (Park 2022). Patients should be informed and educated, e.g., by the physicians, and understand how comorbidities, lifestyle behavior, and modifiable factors affect their cancer risk.
As mentioned above, the comorbidity number was related to the cancer presence. The most common comorbidities were hypertension, obesity, diabetes, and cardiovascular diseases (Furau et al. 2021b, a). Multiple comorbidities, specifically those related to the metabolic syndrome, were also found to be more prevalent, e.g., in uterine cancer patients than in the general population, and this difference persisted after adjustment for age (Cook et al. 2013). Thus, we concluded, similarly to other authors, that medical programs should be undertaken toward treating comorbidities in EC to improve health quality and prolong survival and recurrence-free survival for these patients (Kurnit et al. 2015; Binder et al. 2016).
Primarily, we analyzed the molecular factors, such as HIF1A, EPAS1, and VEGFA gene expression levels, that could influence cancer risk and development. However, since the abovementioned age, BMI, and comorbidity number differed between cancer patients and the control group, we investigated if there is a relation and/or disturbances between those parameters and expression for the GOIs in these groups. Only EPAS1 was weakly correlated with age in controls but not cancer patients. It seems that the analyzed GOIs expression levels are not affected by age, BMI, and comorbidities.
We analyzed the GOIs expression level and established differences between tissue specimens obtained from cancer-affected and control patients. HIF1A and VEGFA expression were significantly higher in cancerous tissue; EPAS1 did not differ between those groups. Our results regarding HIF1A stay in line with the observations of other researchers. A meta-analysis of 25 studies showed positive associations of HIF1A protein overexpression with tumor grade, lymph node invasion, and invasion of the cancer-affected cells into the myometrium. It also seems to be a poor prognosis predictor in the EC (Zhu et al. 2020). In other gynecological malignancies—ovarian cancer, we also observed differences in HIF1A and VEGFA expression levels in a case–control study, but also EPAS1 differed significantly (Englert-Golon et al. 2022). Similarly to the results shown in this study, the VEGFA expression was higher in cancer-affected tissue, whereas the HIF1A protein was at a lower level in malignant ovarian tumors. Thus mutual relationships in the expression of both HIF1A and EPAS1 and VEGFA but also with other genes and proteins could be of importance also in endometrial cancer progression (Englert-Golon et al. 2022). As HIF1A plays an essential role in the adaptive cellular response to hypoxia and is associated with poor clinical outcomes, it seems to be a promising therapeutic target in the EC course of treatment. Thus, several selective and non-selective inhibitors directly or indirectly target the HIF1A upstream or downstream signaling pathway and, as a result, decrease this protein level (Seeber et al. 2010).
Moreover, previous studies showed that the modulation of hypoxia-inducible factors and vascular endothelial growth factor A might contribute to the activation of downstream signaling pathways involved in cancer progression and related processes such as neovascularization (D’amico et al. 2021). As a result, hypoxia-induced angiogenesis triggers the epithelial-mesenchymal transition process (Maugeri et al. 2016, 2021). Additionally, mutual relationships of both HIF1A and EPAS1 with VEGFA play an important role in the oncogenesis and progression of other gynecological malignancies (Englert-Golon et al. 2022). Over the past years, the proteins HIF1A (Semenza 2000; Melillo 2006), EPAS1 (Petrella and Brinckerhoff 2009; Wigerup et al. 2016; Singhal et al. 2019), and VEGFA (Terme et al. 2013; English et al. 2017; Yang et al. 2018) have been shown to be eligible targets for anti-tumor therapies.
We observed significantly higher expression levels of VEGFA in endometrial epithelial tumors (Dziobek et al. 2019). VEGFA was shown to be regulated together with HIF1A by dipeptidyl peptidase IV, which plays a direct role in the progression of several human malignancies (Khin et al. 2003; Beckenkamp et al. 2015; Yang et al. 2017). VEGFA, with VEGFB and their receptors, are crucial proteins involved in the development of new blood vessels. If the blood flow is suboptimal, its expression depends on a hypoxia environment (Dziobek et al. 2019). We observed stronger relations between the gene-to-gene expression level in tissues that lack cancerous changes (correlation coefficients 0.56–0.66) compared to neoplasia-affected tissues (correlation coefficients 0.34–0.45). Similar observations were made in the case of ovarian malignancies, where in non-affected tissue, the expression levels of all analyzed genes were significantly and more strongly correlated than in benign changes and malignant tumors (Englert-Golon et al. 2022). It could imply that mutual relations of the analyzed genes or disturbances in their expression control are either a cause or a reason for EC manifestation. However, it was shown that comparing HIF1A to EPAS1 proteins, despite their primary role as transcription factors for cellular response to hypoxia, could play independent or/and coregulatory roles in tumor physiology and progression (Davis et al. 2022; Song et al. 2022).
VEGFA showed the highest dependence of all analyzed genes with tumor grading. We established significant differences between control and tumor grades G2 and G3 and a significant increasing trend in the growth of the VEGFA expression level with the grade. The same tendency was shown in ovarian tumors, and VEGFA expression intensified with the pathological process progress (Englert-Golon et al. 2022). Similarly to VEGFA, regarding HIF1A in endometrial tissue, a significant trend was observed with increasing grading but not in the case of EPAS1. However, we did not observe differences in the expression levels for either HIF1A or EPAS1. We also observed a significant correlation between both HIF1A and VEGFA expression levels and tumor grading but not in the case of EPAS1. It was estimated that HIF1A proteins positively correlated with poor prognosis in EC (Seeber et al. 2010), and it was also shown that VEGFA factors significantly differed between tumor grading (Dziobek et al. 2019). Both are fundamental proteins involved in the hypoxia-vascularization axis and properly nourish and oxygenate non-pathological and cancer cells (El-Sayed Mohammed Youssef et al. 2015; Dziobek et al. 2019). The increase in HIF1A and VEGFA factors was shown to be significantly, directly, and positively associated with increased malignancy in other types of cancer, e.g., oral squamous cell carcinoma (El-Sayed Mohammed Youssef et al. 2015; Mahecha and Wang 2017). Thus, similarly to HIF1A, at least the VEGFA gene could be a promising target for anticancer therapy or promote the efficacy of different treatments (Song et al. 2022).
Menopausal status and GOIs expressionThe additional analyzed parameter was the menopausal status of the patients. Among analyzed GOIs, only the VEGFA expression level was significantly higher in tissue obtained from post-menopausal women compared to pre-menopausal women. However, in the case of gene-to-gene correlations, stronger associations were observed in pre-menopausal women (correlation coefficients 0.72–0.75 vs. 0.27–0.33, respectively). However, it could also be a combined effect of the menopausal and cancer-presence-related status described above. At the same time, we observed a significantly higher percentage of cancer manifested in women after menopause. We suspect, and it could not be excluded, that the effect of menopausal status and cancer manifestation on GOIs expression is mutual in the case of these patients. Thus, we also examined the gene expression differences in case–control studies between pre- and post-menopausal cases. HIF1A and VEGFA expression levels in pre-menopausal controls were lower than in pre-menopausal EC cases. In cancerous tissue samples obtained from post-menopausal subjects, the VEGFA level was higher compared to controls. In turn, the EPAS1 level was higher in non-affected controls before menopause.
Interestingly, in non-cancer-affected controls, all gene-to-gene correlations were positive and strong in pre-menopausal cases and were either moderate for VEGFA and EPAS1 or not significant for other gene-to-gene correlations in post-menopausal. In cancer-affected tissue, significant correlations of VEGFA with HIF1A and EPAS1 were weak and only in post-menopausal cases. These results suggest that in cancer cases and especially after menopause, the mutual relations between analyzed genes weaken and pass over control mechanisms disturbing angiogenesis and the hypoxia balance. As a result, it explains that this process leads to tissue abnormalities. There is still not a very clear understanding of the association between age, menopause, and EC (Wu et al. 2019). Therefore, it is important to analyze other genetic and epigenetic factors that could be allied with cancer susceptibility and progression. Our investigation follows the need to examine the modifiable and non-modifiable factors related to the patient’s condition and the association of HIF1A, EPAS1, and VEGFA with EC risk and progression.
Multivariable adjustment analysesWe conducted multiple linear regression analyses as many factors could influence cancer manifestation. The most significant contributing factors to EC were menopause status, increasing HIF1A levels, and a high BMI. Age was excluded as a redundant factor to the menopause value. Interestingly, EPAS1 expression was the first under the cut-off line and, therefore, would not exclude this gene as being of minor importance in the susceptibility to cancer.
Additionally, we adjusted the analyzed cases to age, BMI, and comorbidity number using multivariable adjustments. After adjustments, the controls and cancer-affected tissue samples showed significantly higher HIF1A expression and slightly, but not significantly, higher VEGFA expression levels. The multivariate analysis showed that classical risk factors are not the only reason for cancer manifestation in patients. To better understand EC biology, we should consider factors leading to changes in gene expression, especially angiogenesis and hypoxia-related genes. The clinical effectiveness of chemotherapy is variable, suggesting that novel molecular targeted therapies against pathways associated with cancer cell survival in EC treatment are needed. As a result, they may impair the cellular processes activated by hypoxia in the tumor microenvironment (Salinas-Vera et al. 2022).
Limitation of the studyThis study also has potential limitations. The groups’ characteristics, in terms of age, BMI, and comorbidities number, differ significantly before adjustment. This issue may influence the presented results, which should be interpreted cautiously. The results are promising, and in-depth analyses, also at protein level in larger sample sizes may validate our observations. Still, as compared to other studies cancer types, and also, e.g., murine models, the analyzed by us genes could deal even as independent prognostic factors and could be potential target for anti-tumor therapies (Joshi et al. 2014; Beuselinck et al. 2018; Wierzbicki et al. 2019; Englert-Golon et al. 2022). Although in this work, we evaluated the mRNA expression of analyzed genes. We are aware that the tissue-specific protein effective translation could differ at some point. However, the effective translation of mRNA to protein was proven in many papers, not only in human endometrial cancer cells (Downes et al. 2018), in endometrial unchanged and cancer-affected specimens (Sivridis et al. 2002; Maybin et al. 2018), but also in other tissues (Maugeri et al. 2016, 2021; Wierzbicki et al. 2019). Additionally, the Human Protein Atlas database showed that most tissues exhibited moderate to strong VEGFA cytoplasmatic staining, weak to moderate HIF1A cytoplasmic and/or nuclear immunoreactivity (The Human Protein Atlas. https://www.proteinatlas.org; Accessed 21 Oct 2022), and other sources confirmed effective translation of EPAS1 gene (Sivridis et al. 2002).
Comments (0)