
Remember me
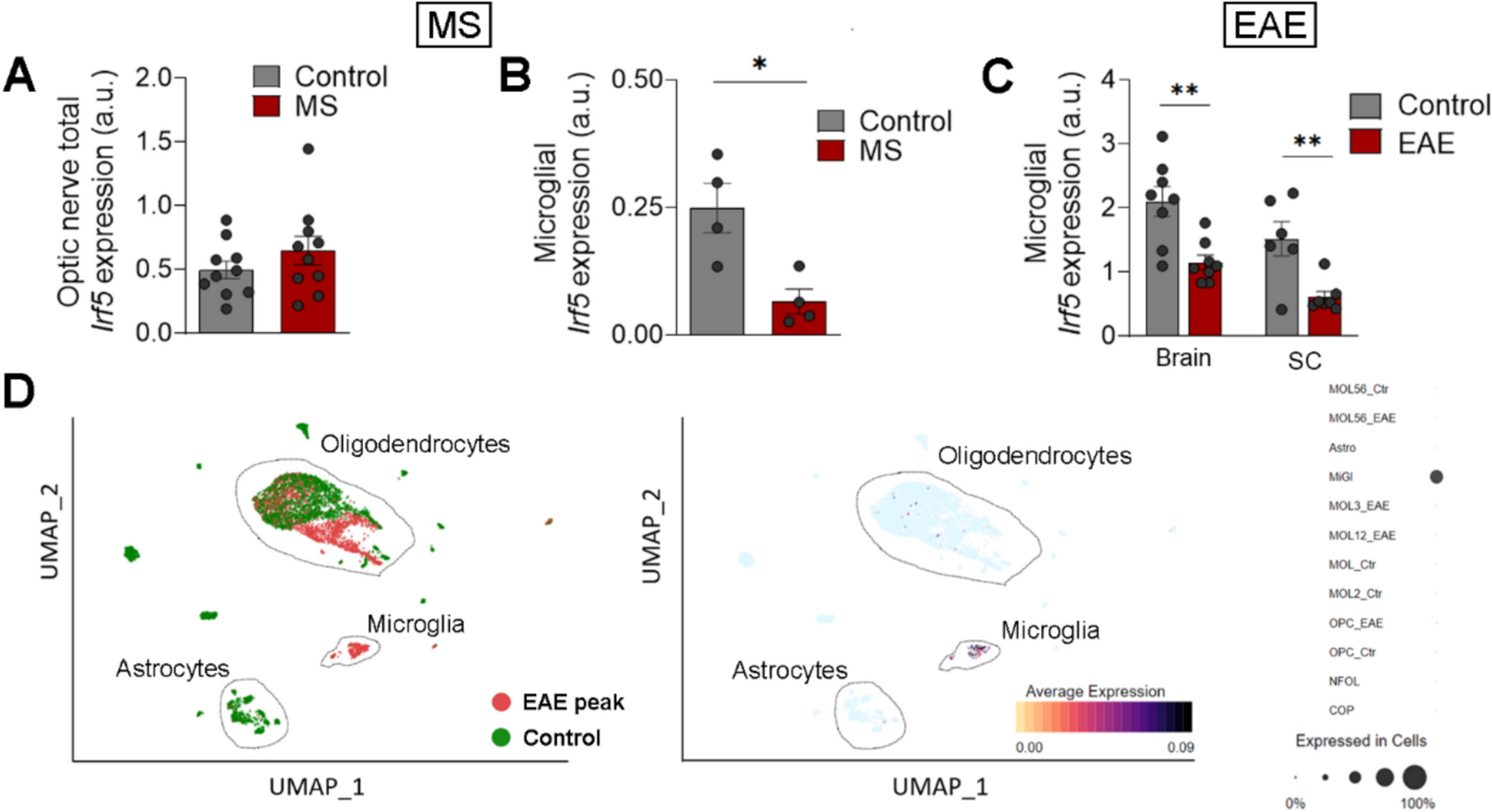


To investigate the effects of dietary Zn supplementation on hepatic Zn metabolism, we determined hepatic Zn content, and the expression of genes related to Zn metabolism. Zn content of liver and hepatic Golgi apparatus increased with dietary Zn levels (Fig. 1a and b). Compared with the L- and M-Zn groups, H-Zn up-regulated the znt6 and mtf-1 mRNA abundance (Fig. 1c and d), and increased the protein levels of ZnT6, total-MTF-1 and nuclear-MTF-1 (Fig. 1e and f). Moreover, immunofluorescence staining corroborated the Zn-dependent upregulation of ZnT6 and MTF-1 at protein levels (Fig. 1g and h). Overall, these results suggested that high dietary Zn led to hepatic Zn overload, increased ZnT6 and MTF-1 expression, and promoted MTF-1 nuclear translocation.
Fig. 1
Effects of dietary Zn levels on Zn metabolism in the liver of yellow catfish. (a), Zn content in the liver. (b), Zn content in the Golgi apparatus of the liver tissue. (c) and (d), mRNA levels of znt6 (c) and mtf-1 (d). (e-f) Western blot (e) and quantification (f) analysis of ZnT6, MTF-1 and n-MTF-1 (nucleus-MTF-1). (g), Immunofluorescent analysis of ZnT6. Scale bars, 75 μm. (h), Immunofluorescent analysis of MTF-1. Scale bars, 25 μm. Values represent the means ± SEM (n = 3 replicate tanks, and 6 fish were sampled from each tank). Significant variations at P < 0.05 are denoted by the Letter (a-c)
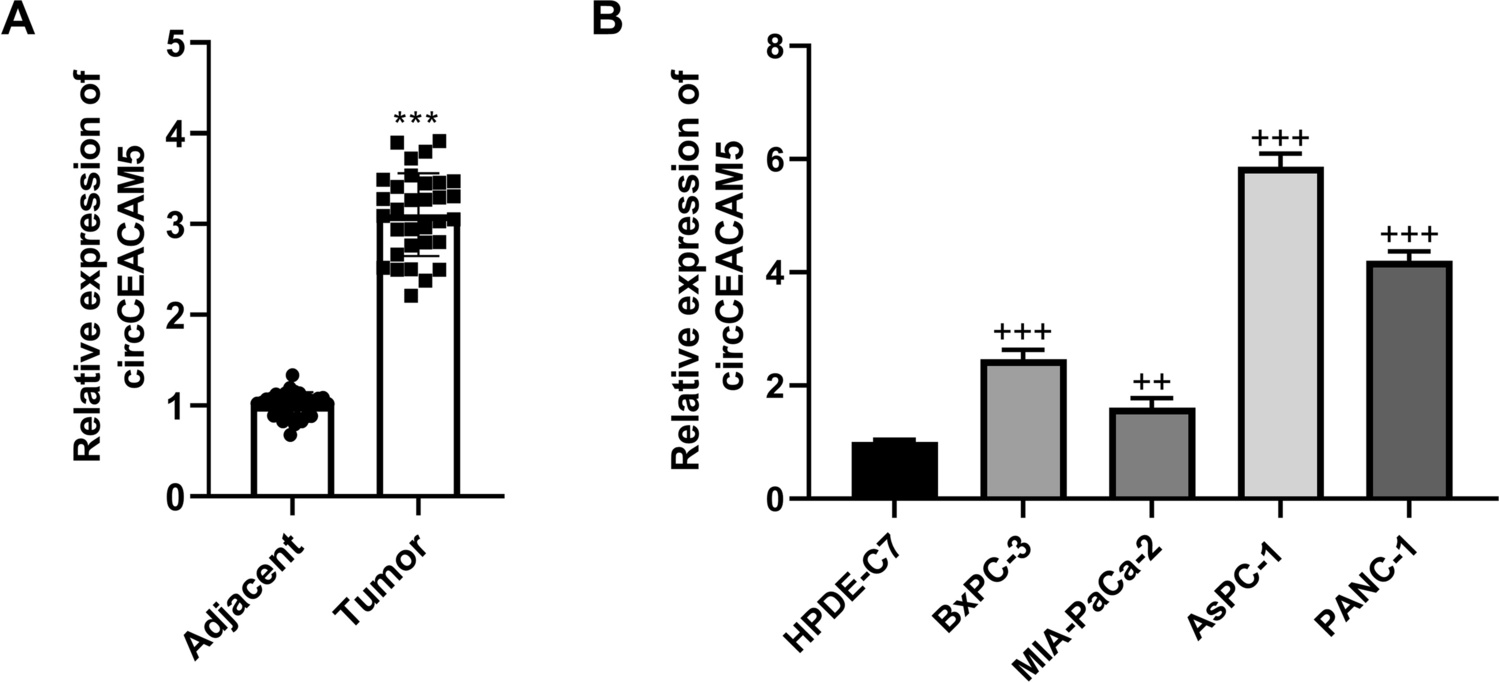
Dietary zn addition influenced SUMOylation progress in the liverSince Zn has been found to influence SUMOylation modification [40], we investigated the effects of dietary Zn supplementation on the expression of genes involved in SUMOylation at the protein and mRNA levels. Our results showed that high dietary Zn reduced SUMO1 protein expression but increased the SUMO-specific proteases (SENP1 and SENP2) protein levels. Among three dietary groups, SUMO2/3 protein expression was highest in M-Zn group, and the lowest in H-Zn group. Compared with the L-Zn group, H-Zn diet significantly reduced the protein levels of E1-activating enzyme SAE1, E2-conjugating enzyme UBC9 and E3 ligase PIAS1, respectively (Fig. 2a and b). Immunofluorescence staining corroborated the Zn-dependent reduction of SUMO1 expression and increase of SENP1 expression (Fig. 2c and d).
Fig. 2
Effects of dietary Zn levels on SUMOylation progress in the liver of yellow catfish. (a and b), Western blot (a) and statistical analysis (b) of the protein involved in pro-SUMOylation (SUMO1, SUMO2/3, SAE1, UBC9 and PIAS1) and de-SUMOylation (SENP1 and SENP2). (c and d), Immunofluorescent analysis of SUMO1 (c) and SENP1 (d). Scale bars, 25 μm. (e), mRNA levels of genes involved in SUMOylation modification. Values represent the means ± SEM (n = 3 replicate tanks, and 6 fish were sampled from each tank). Significant variations at P < 0.05 are denoted by the letter (a-c)
At the transcriptional level, H-Zn diet down-regulated the sumo1, sae1, ubc9 and pias1 mRNA levels, but up-regulated the senp2 and senp6 mRNA expression, compared to the other two groups. Among three dietary groups, sumo3 mRNA expression was highest, and senp3 mRNA expression was lowest for fish fed the M-Zn diet. The mRNA expression of senp1 was significantly lower in L-Zn group than that in other two groups. The sumo2, senp5, senp7 and senp8 mRNA expression was not significantly affected by dietary Zn supplementation (Fig. 2e). Overall, these results indicated that high dietary Zn addition inhibited SUMOylation modification through increasing the expression of SUMO-specific proteases.
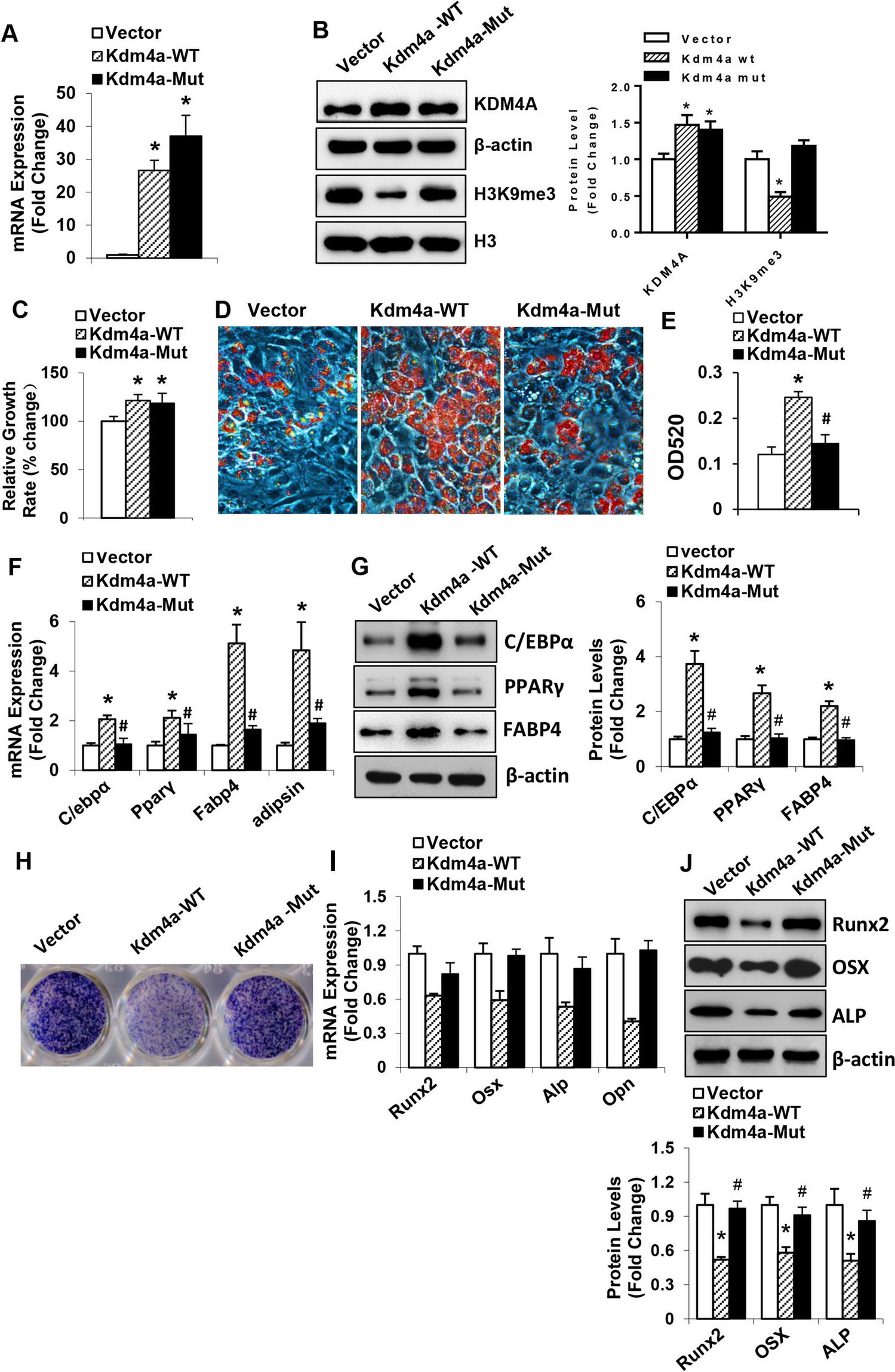
In vitro studiesZn affected Zn metabolism, induced intracellular zn deposition and modulated SUMOylation modification in yellow catfish hepatocytesTo further investigated the effect of Zn incubation on Zn metabolism, as well as the expression of SUMOylation-related genes, hepatocytes were isolated from yellow catfish, and TPEN was used to chelate the Zn. The CCK-8 assay demonstrated that 0–70 µM Zn did not remarkably affect the viability of yellow catfish hepatocytes (Fig. S1A). In order to explore whether the addition of Zn and TPEN affects other transition elements, the contents of Zn, Fe, Cu and Mn in cells were detected. TPEN could not only reduce the intracellular Zn content, but also alleviate the increase of intracellular Zn content induced by excessive Zn addition (Fig. S1B). However, the addition of Zn or TPEN had no significant effect on the contents of other transition elements in the cells, including Fe, Cu and Mn (Fig. S1C-E). Furthermore, TPEN pre-incubation alleviated the Zn-induced increase in ZnT6 protein levels (Fig. 3a and b). Compared with control group, Zn increased znt6 mRNA expression, while TPEN pretreatment significantly alleviated the changes induced by Zn (Fig. 3c). Considering that ZnT5 can form a heterodimer with ZnT6, we also detected the protein expression of ZnT5. Our result indicated that TPEN treatment alleviated the increase of ZnT5 protein expression induced by Zn addition (Fig. S1F and G). Moreover, TPEN pre-incubation remarkably blunted the Zn-induced accumulation of Zn in the Golgi apparatus (Fig. 3d). Using the Newport green DCF Diacetate (N7991) probe, we found that 70 µM Zn treatment increased intracellular Zn content and its co-localization with the Golgi apparatus, while TPEN pre-treatment abrogated these changes (Fig. 3e). Thus, Zn supplementation promoted the transport of Zn into Golgi apparatus.
Fig. 3
TPEN attenuated Zn-induced increase in ZnT6 expression, decrease of SUMO level and the colocalization of ZnT6 and SENP1 in the hepatocytes of yellow catfish. (a and b), Western blot (a) and statistical analysis (b) of ZnT6. (c), mRNA level of znt6. (d), Zn content in Golgi apparatus. (e), Confocal microscopic image of Zn ions (green) co-localized with Golgi (red) by immunofluorescence staining. Scale bars, 25 μm. (f and g), Western blot and statistical analysis of the protein involved in pro-SUMOylation (SUMO1, SUMO2/3, SAE1, UBC9 and PIAS1) and de-SUMOylation (SENP1). (h), Confocal microscopy image of ZnT6 co-localized with SENP1 in yellow catfish hepatocytes detected by immunofluorescence staining using anti-ZnT6 (green) and anti- SENP1 antibodies (red). Scale bars, 4 μm. Values represent the means ± SEM (n = 3 independently biological experiments). The statistical significance (P value) was determined by Student’s t test. *P < 0.05
Compared with the control, Zn incubation down-regulated the protein levels of SUMO1, SUMO2/3, SAE1, UBC9 and PIAS1, but up-regulated the protein expression of SENP1, and TPEN pretreatment alleviated these changes in protein levels induced by Zn (Fig. 3f and g). Furthermore, pre-incubation with TPEN remarkably inhibited Zn-induced increase in the ZnT6 and SENP1 protein levels, and decreased the co-localization of ZnT6 with SENP1, which indicated that Zn incubation potentially promoted the deSUMOylation of ZnT6 by SENP1 (Fig. 3h). Compared to the control, Zn incubation increased the mRNA levels of senp1, senp2, senp6 and senp8, and decreased the mRNA abundance of sumo1, sumo3, sae1, ubc9 and pias1, while these changes were abolished by TPEN pre-incubation (Fig. S1H). Collectively, the above findings suggested that Zn treatment affected the absorption and transport of Zn, led to accumulation of Zn in Golgi apparatus, and modulated SUMOylation modification.
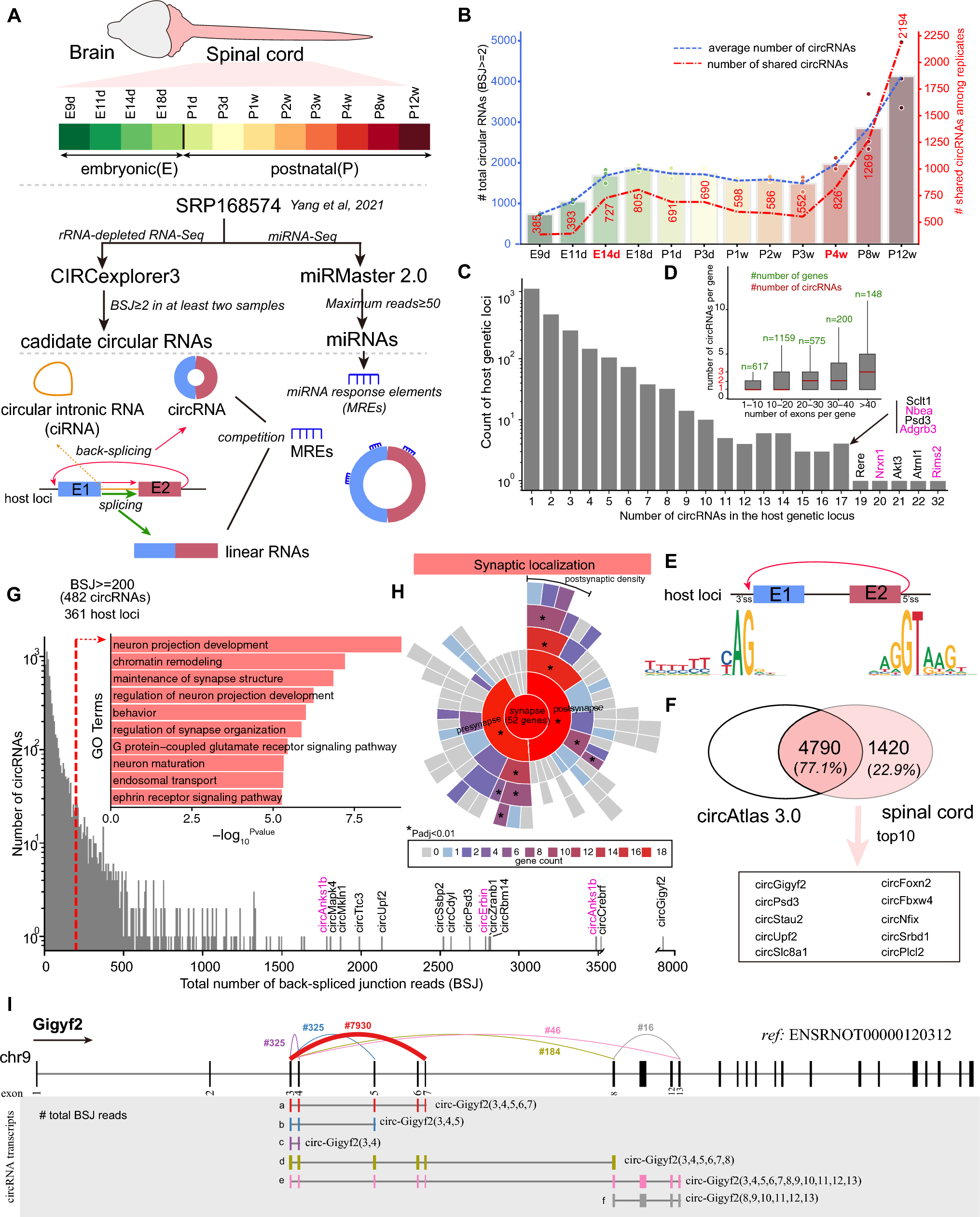
Zn regulated senp1 transcription via MTF-1 binding to its promoterConsidering that high dietary Zn up-regulated the mRNA and protein levels of mtf-1 and senp1, and also promoted translocation of MTF-1 to the nucleus, we conducted in vitro studies to determine whether MTF-1 mediates Zn-induced increase of senp1 expression. TPEN pretreatment significantly alleviated Zn-induced increase of mtf-1 mRNA expression (Fig. S2), total-MTF-1 and nuclear MTF-1 protein levels (Fig. 4a-b) in yellow catfish hepatocytes. Immunofluorescence analysis further confirmed these results (Fig. 4c-d).
Fig. 4
Zn induced the upregulation of SENP1 expression by promoting the DNA binding of MTF-1 to the senp1 promoter. (a-b), Western blot and statistical analysis of MTF-1 and n-MTF-1 (nucleus-MTF-1) in the hepatocytes of yellow catfish. (c-d), Immunofluorescent analysis of MTF-1 (d) and quantification of the co-localization between MTF-1 and nuclear (c). Scale bars, 10 μm. (e), MTF-1 binding sequence (MRE) located at − 589 bp to − 602 bp of senp1 promoter of yellow catfish. (f), Site mutagenesis of MTF-1 on the pGl3- senp1 -951/+99 vector. (g and h), EMSA analysis of predicted MTF-1 binding sequences (MRE) on the senp1 promoter. The 5′-biotin labeled double-stranded oligomers were incubated with nuclear protein in HEK-293T cell (g) or yellow catfish hepatocytes (h). Values represent the means ± SEM (n = 3 independently biological experiments). Asterisk (*) and hash sign (#) represent significant differences at P < 0.05 between the two groups, as determined using Student’s t-test
Next, we performed promoter analysis of senp1 gene to explore whether Zn-induced changes in Zn transporters expression was mediated by MTF-1 targeting the senp1 gene. By analyzing the senp1 promoter (Fig. S3), we predicted an evolutionary conserved MTF-1 binding sequence (MRE), which was located at − 589 to − 602 bp on yellow catfish senp1 promoter (Fig. 4e). Furthermore, Zn treatment increased the luciferase activity of the senp1 promoter (Fig. 4f), and these changes induced by Zn were alleviated by the MRE mutation on senp1, demonstrating that Zn triggered the activation of MTF-1, which in turn transcriptionally activated senp1 via binding of MTF-1 to the MRE motif of the senp1 promoter (Fig. 4f). Meanwhile, EMSA indicated that the putative MRE sequence of the senp1 promoter could bind to nuclear extract, and the interaction could be disrupted by unlabeled probes but not by mutant probes (Fig. 4g and h). In addition, the binding activity of MTF-1 to MRE was significantly enhanced upon Zn incubation (Fig. 4g and h, lane 5), indicating that the − 589 to − 602 bp region of the senp1 promoter was the functional site for its transcriptional regulation.
Then, small interference RNA si-mtf1 was used to determine the correlation between MTF-1 and SENP1 in yellow catfish hepatocytes. According to mRNA and western blotting analysis, siMTF-1-2 had the highest knockdown efficiency and was selected for our experiment (Fig. S4A-C). The siMTF-1 pretreatment significantly alleviated the Zn-induced up-regulation of the SENP1 mRNA and protein levels (Fig. S4D-F). Overall, the above results demonstrated that the MRE mediated the transcriptional activation of senp1 to Zn overload.
SENP1 mediated Zn-induced upregulation of the mRNA and protein levels of ZnT6Having obtained strong evidence that Zn overload promoted SUMO-specific proteases expression and affected the expression of ZnT6, we further investigated the potential causal relationship between them via the small interference RNA (siRNA) against SENP1 expression. siSENP1-1 had the highest knockdown efficiency based on the results of mRNA and western blotting analysis (Fig. 5a-c). Immunofluorescence staining further corroborated that siSENP1-1 had the highest interference efficiency (Fig. 5d). Therefore, siSENP1-1 was selected for subsequent experiments. The senp1 knockdown suppressed the Zn-induced up-regulation of SENP1 protein abundance (Fig. 5e and f). Furthermore, si-SENP1 treatment attenuated the increase of ZnT6 protein abundance induced by Zn (Fig. 5e and g). Collectively, these data indicated that Zn-induced increase of ZnT6 expression was mediated by SENP1.
Fig. 5
The SENP1 mediated Zn-induced increase of ZnT6 expression in the hepatocytes of yellow catfish. (a-c), qPCR (a), Western blot and statistical analysis (b and c) of the knockdown efficiency of siRNA targeting senp1 in hepatocytes. (d), Confocal microscopic image of the SENP1 protein detected by immunofluorescence staining. Scale bars, 4 μm. (e-g), Western blot and statistical analysis of SENP1 (e and f) and ZnT6 (e and g) protein expression. Values represent the means ± SEM (n = 3 independently biological experiments). The statistical significance (P value) was determined by Student’s t test. *P < 0.05
ZnT6 was SUMOylated by SUMO1 and deSUMOylated by SENP1To identify whether ZnT6 could be SUMOylated in cells and which type of SUMO could be conjugated with ZnT6, we transiently transfected HA-ZnT6 with the different types of SUMO (Myc-tagged SUMO1/2/3) into HEK-293T cells. Then, immunoprecipitation (IP) was conducted to determined SUMOylation of ZnT6. Cell lysates were collected and immunoprecipitated with anti-HA antibody. Anti-HA immunoblotting results showed a specific shifted band of SUMOylated-ZnT6. According to the gray value of the SUMOylated-ZnT6 band, we found that ZnT6 was modified mainly by SUMO1 (Fig. 6a). To further confirm the role of SUMO1 in the SUMOylation of ZnT6, we constructed a mutant SUMO1 by deleting the diglycine residues form the thioester linkage. Our results showed that ZnT6 SUMOylation can be detected only when ZnT6 is co-transfected with wild-type SUMO1 in HEK-293T cells, while SUMO1 mutant cannot (Fig. 6b). Our further immunofluorescence analysis revealed the co-localization between ZnT6 and SUMO1 (Fig. 6c). Therefore, we mainly focused on SUMO1-mediated SUMOylation of ZnT6 in the following experiments. To further verify whether the only E2-conjugating enzyme (UBC9) participated in the SUMOylation of ZnT6, we transfected HA-ZnT6 alone or in combination with Myc-tagged SUMO1, His-UBC9 into HEK-293T cells. We found that UBC9 overexpression significantly enhanced the SUMOylation of ZnT6, indicating the important role of UBC9 in ZnT6 SUMOylation modification (Fig. 6d, lanes 4). Co-immunoprecipitation (Co-IP) analysis revealed that HA-ZnT6 interacted with His-UBC9, indicating their physical link in HEK-293T cells (Fig. 6e and f). These results supported that ZnT6 was directly conjugated by SUMO1 in human cells.
Fig. 6
ZnT6 was mainly SUMOylated by SUMO1 and deSUMOylated by SENP1. (a), ZnT6 was mainly modified by SUMO1. HA-ZnT6 and Myc-SUMO1/2/3 were transfected into HEK-293T cells. The cell lysates were then IP with anti-HA antibody, followed by immunoblotting with the identical antibody. (b), Immunoprecipitation and immunoblot analysis of HEK-293T cells co-transfected with expressing vectors of HA-ZnT6, His-UBC9, Myc-SUMO1 and its indicated mutants. (c), Confocal microscopy image of ZnT6 co-localized with SUMO1 in yellow catfish hepatocytes detected by immunofluorescence staining using anti-ZnT6 (green) and anti-SUMO1 antibodies (red). Nuclear were stained blue. Scale bars, 4 μm. (d), UBC9 enhanced SUMOylation of ZnT6. HEK-293T cells were co-transfected with HA-ZnT6, Myc-SUMO1 and with or without His-UBC9, SUMOylated bands were then detected with anti-HA or anti-Myc antibody. (e and f), The interaction between ZnT6 and UBC9. The lysates of HEK-293T cells overexpression HA-ZnT6 and His-UBC9 were processed for IP with anti-HA (e) or anti-His (f) antibody, followed by immunoblot analysis with the reciprocal antibody. (g), SENP1, but not SENP2, 3, 5, 6, 7 and 8, deSUMOylated ZnT6. HA-ZnT6, Myc-SUMO1 and indicated isoforms of SENPs were transfected into HEK-293T cells, and then western blotting was performed to analyze the SUMOylation of ZnT6 with anti-HA antibody. (h), The deSUMOylated of ZnT6 is dependent on the catalytic activity of SENP1. HA-ZnT6, Myc-SUMO1 and His-UBC9 were transfected with wild-type SENP1 or the SENP1 C697A mutant into HEK-293T cells as indicated. Cell lysates were subjected to IP with anti-HA antibody, followed by immunoblot analysis using anti-HA antibody. (i), SENP1 down-regulated ZnT6 SUMOylation in a dose-dependent manner. HA-ZnT6 and Myc-SUMO1 were transfected into HEK-293T cells with varying amounts of Flag-SENP1, and then western blotting was performed to analyze the SUMOylation of ZnT6 using specified antibodies. (j), Zn incubation prevented ZnT6 from SUMOylation modification in HEK-293T cells. HA-ZnT6, Myc-SUMO1 and His-UBC9 were transfected into HEK-293T cells. After treatment with or without Zn (70 µM) for 24 h, the cell lysates were subjected to IP with anti-HA antibody, followed by immunoblotting with the same antibody to detect the SUMOylation of ZnT6 under Zn treatment. The band representing SUMOylation of ZnT6 was labeled with an arrow
SUMOylation is a dynamic process which was reversed by SUMO-specific proteases (SENPs), collectively known as deSUMOylase [22]. According to the released genome sequence, seven SENPs have been identified in yellow catfish (SENP1, 2, 3, 5, 6, 7 and 8) [39]. To identify which SENP removed the SUMO1 modification, SUMO1 and each of the seven SENPs were co-transfected with HA-ZnT6 into HEK-293T cells, and then SUMOylation of ZnT6 was determined. The results showed that SENP1 reduced ZnT6 SUMOylation level, while other SENPs did not influence ZnT6 SUMOylation level (Fig. 6g). In mammals, the C603A mutant of SENP1 did not have catalytic activity [41]. Importantly, sequence alignment found that C697 site of yellow catfish SENP1 (corresponding to C603 in mammalian SENP1) appears to be highly conservative among different species (Fig. S5). Thus, the overexpression vector of SENP1C697A was generated from the full-length SENP1, which contains the mutant form of C697A (cystine was mutated to alanine at 697 position). We found that the wild-type SENP1 reduced the SUMOylation level of ZnT6, while the SENP1C697A did not (Fig. 6h). The SUMOylation level of ZnT6 appeared to be decreased in a dose-dependent manner in the case of SENP1 overexpression (Fig. 6i), further confirming the critical role of SENP1 in mediating ZnT6 deSUMOylating. Furthermore, to determine whether Zn affected SUMOylation modification of ZnT6, HA-ZnT6, Myc-SUMO1 and His-UBC9 were co-transfected into HEK-293T cells and treated with or without indicated concentrations of Zn for 24 h, showing that Zn treatment greatly inhibited SUMOylation modification of ZnT6 (Fig. 6j). All above results proved that SENP1 was a crucial deSUMOylation enzyme for ZnT6 in human HEK-293T cells.
Lysine 409 (K409) was the major SUMOylation site of ZnT6Based on the SUMOplot™ Analysis Program (http://www.abgent.com/sumoplot), the potential SUMOylation sites of ZnT6 were predicted, which were located on multiple lysine residues at amino acid positions K29, K47, K162 and K409 in yellow catfish. (Fig. 7a). To determine which of these lysine residues was the most important for SUMOylation of ZnT6 in yellow catfish, the Lys residues (K) were individually replaced with Arg residues (R). Then the plasmids expressing SUMO1 and UBC9 were co-transfected with WT or the mutant ZnT6 constructs into HEK-293T cells, and the levels of SUMOylation were analyzed as performed previously. Compared with wild-type ZnT6, mutants K29R, K47R and K162R did not cause significant changes in the SUMOylation levels of ZnT6 (Fig. 7b, lanes 4–6), and only the K409R mutant dramatically decreased ZnT6 SUMOylation level (Fig. 7b, lane 7), indicating that K409 of ZnT6 was the major SUMOylation site for SUMO1.
Fig. 7
Lysine 409 (K409) is the major SUMOylation site of ZnT6. (a), Prediction of four potential SUMOylation sites of ZnT6 in yellow catfish using SUMOplot software. (b), K409 was the major SUMOylation site in ZnT6. The lysates from HEK-293T cells overexpressing HA-ZnT6, HA-ZnT6 mutants (K29R/K47R/K162R/K409R), Myc-SUMO1 and His-UBC9 were IP with anti-HA antibody, and then immunoblot analysis was performed with the same antibody. (c), The mutant ZnT6-F408A/D410E further confirmed the SUMOylation of ZnT6. The lysates from HEK-293T cells overexpressing the indicated plasmids were IP with anti-HA antibody, and then immunoblot analysis was performed with the same antibody. (d), Sequences Alignment of ZnT6 from indicated species. The conserved lysine residue (K409) of SUMOylation modification is highlighted in yellow. (e), SUMOylation sites of yellow catfish ZnT6
To further determine the role of lysine 409 in mediating ZnT6 SUMOylation modification. We replaced the two amino acid residues (phenylalanine and aspartic acid) on the flanking region of K409 with alanine to interrupt the consensus sequence of SUMOylation according to previous study [41]. Compared to the WT HA-ZnT6, HA-ZnT6-F408A/D410E co-expressed with Myc-SUMO1 significantly reduced SUMOylation levels of ZnT6, similar to HA-ZnT6-K409R (Fig. 7c). Importantly, sequence alignment revealed that K409 site of ZnT6 in yellow catfish is highly conserved in most of vertebrates (Fig. 7d). The protein structures of human ZnT6 were modeled in I-TASSER server (http://zhanglab.ccmb.med.umich.edu/I-TASSER/), and the structure of chitosan was downloaded from “PUB CHEM” database. The SUMOylation site of ZnT6 was picked up using the online tool pymol (https://sumo.biocuckoo.cn/) (Fig. 7e). Collectively, our results confirmed that K409 was the major site for SUMOylation modification of ZnT6 in yellow catfish.
ZnT6 SUMOylation did not affect its localization, but reduced its protein stabilitySUMOylation participates in various signaling pathways and cellular processes by regulating the stability, subcellular location, transcriptional activity and interaction of targeted proteins [17,18,19]. To check whether SUMOylation modification affected the stability of ZnT6, the CHX assay was performed. HA-ZnT6-WT or HA-ZnT6-K409R was transfected into HEK-293T cells, followed by treatment with the protein translation inhibitor cycloheximide (CHX), for indicated time (0, 3, 6, 9 and 12 h). We found that the half-life of ZnT6-WT was shorter than that of the K409R mutation after CHX incubation (Fig. 8a). In addition, we also tested whether the stability of endogenous ZnT6 is affected by SUMOylation. SENP1 was knocked down by RNAi in yellow catfish hepatocytes. Knockdown of senp1 remarkably decreased the half-life of endogenous ZnT6 (Fig. 8b). Taken together, our study confirmed that the protein stability of ZnT6 could be affected by the SUMO modification.
Fig. 8
SUMOylation of ZnT6 did not affect its localization, but reduced its protein stability, inhibited the increase of Zn content in Golgi apparatus induced by Zn. (a), Western blotting analysis was performed to determine the half-life of ZnT6 by using anti-HA antibody in HEK-293T cells overexpressing wild-type ZnT6 or the ZnT6-K409R with or without CHX incubation for different time. (b), Western blotting analysis was performed to determine the half-life of endogenous ZnT6 by using anti-ZnT6 antibody in yellow catfish hepatocytes treatment with SENP1 knockdown with or without CHX incubation for different time. (c and d), ZnT6 SUMOylation did not change its subcellular localization. (c) Subcellular localizations of nuclear (blue), ZnT6 (green) with Golgi (red) were determined by confocal microscopy. Scale bars, 4 μm. (d) HEK-293T cells overexpressing wild-type ZnT6 or the ZnT6-K409R mutant were immunostained with ZnT6 (green), Golgi (red) and Hoechst (blue). Scale bars, 20 μm. (e and f), ZnT6 SUMOylation inhibited Zn-induced increases of the Zn content in Golgi apparatus. (e) Subcellular localizations of nuclear (blue), Zn ions (green) and Golgi (red) were determined by confocal microscopy. Scale bars, 20 μm. (f) Zn content in Golgi apparatus. Values represent the means ± SEM (n = 3 independently biolological experiments). The statistical significance (P value) was determined by Student’s t test. *P < 0.05
Next, we attempted to investigate whether the subcellular localization of ZnT6 was affected by SUMOylation. Immunofluorescence analysis confirmed that Zn incubation remarkably enhanced the protein expression of ZnT6 and its co-localization with the Golgi apparatus, while siSENP1 pretreatment alleviated this trend (Fig. 8c). At the same time, ZnT6 was mainly localized in the Golgi apparatus, and did not relocate under different levels of SUMOylation (Fig. 8c). In addition, compared with HA-ZnT6-WT, HA-ZnT6-K409R mutant did not significantly alter the subcellular localization of ZnT6 (Fig. 8d). Accordingly, our results demonstrated that ZnT6 SUMOylation did not alter its localization.
ZnT6 SUMOylation inhibited the increase of Zn content in Golgi apparatus induced by ZnOur results above indicated that Zn reduced the SUMOylation level of ZnT6 by increasing the expression of SENP1. To further investigated the effect of ZnT6 SUMOylation level on Zn homeostasis in Golgi apparatus, SENP1 was knocked down by RNAi in yellow catfish hepatocytes. Immunofluorescence analysis indicated that Zn treatment obviously increased the Zn content and its co-localization with the Golgi apparatus, while siSENP1 pretreatment alleviated this trend (Fig. 8e). In addition, the Zn content in Golgi apparatus was determined by ICP-OES. Knockdown of senp1 remarkably alleviated Zn-induced increase of Zn content in Golgi apparatus (Fig. 8f). Taken together, SUMOylation of ZnT6 inhibited the increase of Zn content in Golgi apparatus induced by Zn.
Comments (0)