
Remember me
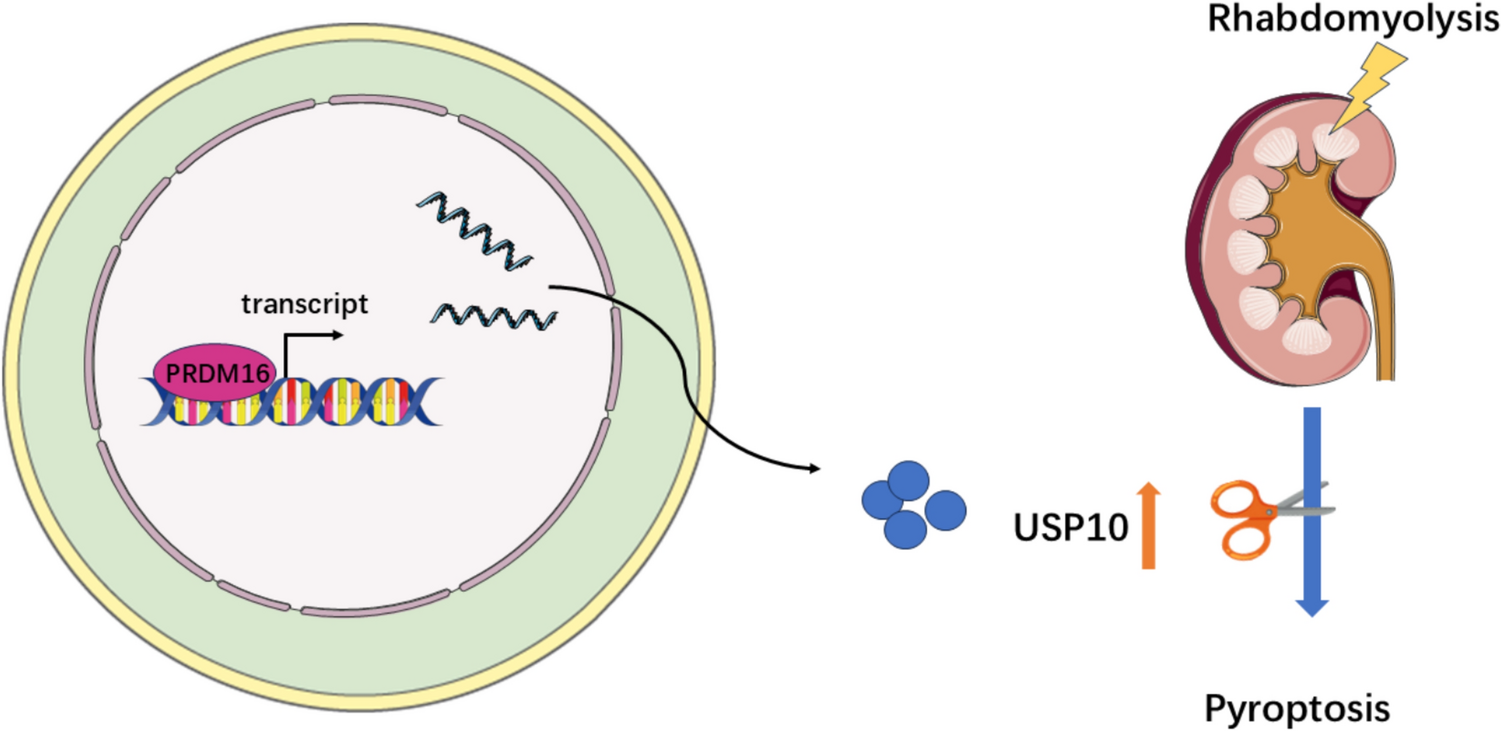


Phospholipid scramblase scramb1 is initially expressed in the subesophageal region of stage 12 embryos, where garland nephrocytes are located. Its expression becomes more robust in later stages of embryogenesis (Fig. 1A and Fig. S1 A). Throughout larval stages, scramb1 continues to be expressed in garland nephrocytes and it is activated in additional tissues, including pericardial cells, the Malpighian tubules and the neuromuscular synapsis (Fig. 1B; Fig. S1 B and [33, 34]).
Fig. 1
scramb1 expression and subcellular localization. (A-B) scramb1 is expressed in the garland nephrocytes of stage 15 Drosophila embryos (A, arrow) and of third instar larvae (B) as detected by in situ hybridization. (C-E’) Distribution of Scramb1-A-V5 driven by the indicated Gal4 lines (anti-V5 antibody) and Duf in garland nephrocytes of third instar (C-D’) and first instar (E-E’) larvae shown at a medial section (C-C’) and at higher magnifications at cortical levels (D-E’). Scramb1-A-V5 colocalizes with Duf in SDs (arrows). Asterisks in C and C’ point to cytoplasmic aggregates. (C) Nuclei were stained with DAPI (blue)
scramb1 is transcribed from two alternative promoters, generating two isoforms, Scramb1-A and -B. Scramb1-A, the predominant isoform in nephrocytes according to RNAseq data (Fig. S1, C and F; and Methods section), is longer due to the inclusion of an N-terminal proline-rich region.
To examine Scramb1 subcellular distribution we generated a V5-tagged version of Scramb1-A and expressed it in nephrocytes. The protein accumulates at the cortical region (Fig. 1, C and C’, arrows), drawing a distinctive fingerprint-like pattern that corresponds to SDs, as revealed by its colocalization with Duf (Fig. 1D-E’, arrows). A cytoplasmic distribution and occasional cytoplasmic aggregates are also observed, possibly caused by non-physiological overexpression conditions (Fig. 1C and C’, asterisks).
Together, these findings indicate that scramb1 is highly expressed in nephrocytes and that the protein localizes within SDs, suggesting it could be a novel component of the SD protein complex.
scramb1 is required for SDs formationTo examine scramb1 function in nephrocytes, we generated novel loss-of-function alleles by the imprecise excision of a P-element inserted in the first intron of the gene (scramb1EY07744). In scramb143, a deletion of 2.9 kb of genomic DNA removes the transcription start sites of all scramb1 isoforms, resulting in a probable null allele. Accordingly, transcripts could not be detected either by RT-PCR or by in situ hybridization (Fig. S1, C-E). scramb143 mutant flies are homozygous viable and fertile, and they do not exhibit any visible macroscopic phenotype, which is consistent with previous reports for another scramb1 allele [33]. In contrast, the larval garland nephrocytes show gross morphological abnormalities. Instead of displaying its distinctive garland-like cellular arrangement, nephrocytes are aggregated in scramb143, a phenotype characteristic of mutations that disrupt SD formation (Fig. 2B, compare with wild-type in A). Accordingly, scramb143 nephrocytes display either no SD strands (25% of nephrocytes) or only a few SD strands (75%) on their surface, mainly near regions of cell contact (n = 110 cells examined; Fig. 2, B and B’, arrows), contrasting with the multitude of SDs that in the wild type describe a dense fingerprint-like pattern on the nephrocyte surface (Fig. 2A’). Interestingly, broad regions of the plasma membrane of mutant nephrocytes are covered by foci of about 400 nm in diameter that accumulate Pyd but not Duf (Fig. 2, B and B’, arrowheads). In addition, Duf and Pyd are coexpressed in some regions of contact between the aggregated nephrocytes (Fig. 2B, asterisk). The described phenotypes are already apparent in first instar larvae, suggesting they are not caused by a possible degeneration of the nephrocytes (Fig. S2 B, compare with wild-type in A). Thus, our data indicate that scramb143 nephrocytes are mostly devoid of SDs.
Fig. 2
scramb1 loss of function phenotype. (A-B’) Immunostaining of wild-type (A-A’) and scramb143 (B-B’) garland nephrocytes depicting the distribution of the SD proteins Duf and Pyd. In scramb143 nephrocytes, SD strands are sparse (arrow, expressing Duf and Pyd). Pyd predominantly accumulates in cortical foci devoid of Duf (arrowheads). Furthermore, Duf and Pyd colocalize in certain regions of contact between clustered nephrocytes (asterisk). (C-C’) Immunostaining of scramb143 larval nephrocytes phenotypically rescued by the expression of UAS-scramb1-A-V5 driven by pros-Gal4, to show the expression of Duf and Pyd, as indicated. SD strands cover the entire nephrocyte surface. (A, B and C) medial planes. (A’, B’ and C’) cortical planes. (D-F) TEM images of scramb143 nephrocytes. An overview of a complete nephrocyte is shown in D (n: nucleus). The highlighted region is shown at higher magnification in E. Electron-dense plaques (black arrowheads in D and E) that bridge the plasma membrane with sub-cortical lacunae are frequently observed. (F) Tangential section through the nephrocyte cortex displaying electron-dense circular structures (black arrowheads) that might correspond to the cortical electron-dense plaques observed in cross-sections. Occasional SDs are also observed (D, red arrows. See also Fig. S2 G). (G) TEM image of a scramb143 mutant nephrocyte rescued by the expression of UAS-scramb1-A-V5, displaying a normal density of SDs (red arrows). Blue arrowheads in E and G point to clathrin coated vesicles and pits. (H) Immunogold labelling of Scramb1-A-V5 (anti-V5 antibody) in a nephrocyte of the same genotype as in G, showing that gold particles associate with SDs (red arrows). Statistical analysis described in the Methods section
The presence of occasional SD-like structures in scramb143 nephrocytes prompted us to examine whether scramb2, a scramb1 paralog enriched in nephrocytes and displaying redundant activity with scramb1 during synaptic transmission (Fig. S1 G, arrow and [33]), may also contribute to SD formation. To this end, we generated novel scramb2 mutations using CRISPR-Cas9 genome-editing technology. scramb2V3, resulting in a frameshift mutation after Asp-16 and truncating 94% of the protein residues (Fig. S1 F), did not exhibit a discernible SD phenotype (Fig. S2, C and C’). In addition, a double mutant scramb143, scramb2V6, an allele that produces a comparable Scramb2 truncation (Fig. S1 F), displayed a nephrocyte phenotype indistinguishable from that of scramb143 (Fig. S2, D and D’, compare with Fig. 2, B and B’). Notably, while the expression of UAS-scramb1-A-V5 in a scramb143 mutant background fully restores SD formation, the expression of UAS-scramb2-HA showed no observable effect (Fig. 2, C and C’; and Fig. S2, E and E’). These findings indicate lack of functional redundancy between the two paralogs in nephrocytes.
As expected from the previous results, examination of the ultrastructure of scramb143 nephrocytes by transmission electron microscopy (TEM) showed an almost complete absence of SDs in the plasma membrane (Fig. 2, D and E; quantitated in Fig. S2 H). This contrasted with the abundant SDs decorating the cortex in the wild-type (Fig. S2 F, arrows; Fig. S2 H; and [10, 11]) and in scramb143 nephrocytes rescued by the expression of UAS-scramb1-A-V5 (Fig. 2G, red arrows; quantitated in Fig. S2 H). Seldom, we observed structures similar to SDs and that probably correspond to the scarce SD-like strands observed by confocal microscopy (Fig. 2D and Fig. S2 G, red arrows; quantitated in Fig. S2 H). A remarkable characteristic of scramb143 mutants is the presence of cisternae that run parallel to the plasma membrane beneath broad regions of electron-dense plaques of about 300–800 nanometers in length and that are absent in the wild-type or in rescued nephrocytes (Fig. 2, D and E, arrowheads; quantitated in Fig. S2 H). Circular electron-dense patches that might correspond to these plaques are occasionally visible in cortical tangential sections (Fig. 2F, arrowheads). These plaques could correspond to the plasma membrane foci that accumulate Pyd in scramb143 nephrocytes observed by confocal microscopy, since their respective sizes and frequencies are compatible (Fig. 2B’, arrowheads). Similarly to the wild-type, abundant clathrin coated vesicles and pits are observed in cell from both scramb143 mutants and rescued animals (Fig. 2, E and G, blue arrowheads), and the remaining ultrastructural organization of the nephrocytes remains largely unaffected.
Next, we examined the distribution of Scramb1-A by immunoelectron microscopy using anti-V5 antibodies in scramb143 nephrocytes rescued by UAS-scramb1-A-V5 expression. To reduce the accumulation of ectopic protein, we switched off UAS-scramb1-A-V5 expression 72 h before fixation. This condition resulted in a complete rescue of SDs (Fig. 2G; quantitated in Fig. S2 H) and in Scramb1-A-V5 mostly located in the cortex, as visualized by confocal microscopy (Fig. S2 I). Notably, Scramb1-A-V5 signal was associated with SDs with a highly statistically significant value (see the Methods section), whereas no other structure was consistently labeled (Fig. 2H, red arrows point to SDs). These results demonstrate that Scramb1 is a constituent of the SD complex in Drosophila and that it is required for its assembly and/or maintenance.
scramb1 is required to recruit Duf to complexes containing Sns, Pyd and Src64BWe have shown that in the absence of scramb1, nephrocytes are largely devoid of SDs. scramb1 might play a role either in the initial assembly of the SD complexes or in their subsequent maturation and maintenance. To better define scramb1 role, we characterized the de novo formation of SDs by inducing the expression of Scramb1-A-V5 in scramb143 nephrocytes during the larval stages using the TARGET technology, which allows the activation of transgenes through a temperature switch [35].
Similarly to scramb143 mutants, at time zero before the expression of Scramb1-A-V5; Pyd and Sns colocalize in foci covering large regions of the nephrocyte plasma membrane (Fig. 3A and Fig. S3, white arrowheads). In contrast, Duf, an essential component of the SDs, is absent from these foci. We interpret these foci as representing aberrant pre-SDs complexes that cannot progress to form SDs in the absence of Scramb1 and in particular, cannot recruit Duf. This suggestion gains support from the observation that the kinase Src64B, the Drosophila ortholog of Fyn involved in SD formation and repair [12], is specifically active within the majority of these foci, as evidenced by phospho-Src64B accumulation (Fig. 3B, white arrowheads).
Fig. 3
Time-course analysis of the induction of SD formation by Scramb1-A. (A-B) Immunostaining of scramb143 or scramb143 nephrocytes rescued by the expression of UAS-scramb1-A-V5 for increasing periods of time (0, 12 and 18 h, as indicated) using the TARGET technology. See the Methods section for the complete genotype. The distribution of Scramb1-A-V5 (anti-V5 antibody), Sns, Pyd, Duf and phospho-Src64B in the cortical region are shown, as indicated. Each image corresponds to a Z-projection of several cortical planes. No SD strands are observed in nephrocytes that do not express UAS-scramb1-A-V5. Instead, abundant cortical foci containing Pyd, Sns and phospho-Src64B cover the nephrocyte surface (white arrowheads). At the 12 h window (two examples shown), Duf is visible in those foci and some acquire an elongated shape (yellow arrowheads). At 18 h, multiple short SD strands cover the surface of the nephrocytes. All images shown at the same magnification. See Fig. S3 for additional time points and medial sections. (C) TEM image of a scramb143 nephrocyte rescued by the expression of UAS-scramb1-A-V5 for 12 h. Multiple SDs sealing small labyrinthine channels are visible (red arrows). Electron-dense plaques, marked by black arrowheads in Fig. 2E, are rare
Scramb1-A-V5 protein becomes detectable in nephrocytes six hours after switching to the permissive temperature. At this time window, the cortical Pyd foci maintain the same morphology but begin to accumulate low levels of Scramb1-A-V5 and Duf (Fig. S3). After six additional hours (12-hour time point), short rods containing Sns, Duf, Pyd and phospho-Src64B can be observed on the surface of nephrocytes (Fig. 3A and Fig. S3, yellow arrowheads), along with circular foci displaying the same set of proteins (Fig. 3A and Fig. S3). Thus, Scramb1-A-V5 is recruited to pre-existing Sns/Pyd/Src64B-containing foci, along with Duf, and is necessary to allow the formation of elongated structures resembling the SD strands observed in the wild-type, albeit shorter. Supporting this interpretation, after 12 h of supplying Scramb1-A-V5, ultrastructural analyses revealed the presence of multiple SDs, usually associated to short labyrinthine channels. Interestingly, the electron-dense plaques characteristic of the scramb143 mutant were almost absent at this time point (Fig. 3C, compare with Fig. 2E). Finally, 18 h after the temperature switch, most nephrocytes were covered by a loose network of strands containing Duf, Sns, Pyd and Scramb1-A-V5, similar to the SD fingerprint pattern observed in the wild-type but less regular and dense, indicating an already significant rescue of the phenotype (Fig. 3A).
These findings suggest that the inability to form SDs in the absence of Scramb1 is due, at least in part, to Duf not being recruited and stabilized in complexes that already contain other essential components of the SD, namely Sns, Pyd and phospho-Src64B. Duf recruitment is likely not mediated through a direct interaction with Scramb1, since Duf is unable to recruit Scramb1 in S2 cells (Fig. S4 A, arrows), indicating the necessity of additional factors absent in this cell line.
The N-terminal proline-rich domain of Scramb1-A protects it from degradation and mediates its association with PydScramb1 localization within SDs could be mediated by its interaction with other SD components. One potential candidate is Pyd, a scaffolding protein that contributes to link the SD complex to the cytoskeleton [36]. Noteworthy, Pyd contains an SH3 domain that could potentially bind to an unstructured, proline-rich region located at the N-terminus of the Scramb1-A isoform (Fig. S1 F). To test this hypothesis, we conducted co-immunoprecipitation assays using extracts from salivary glands expressing both proteins. Since the Pyd isoform expressed in nephrocytes, Pyd-P, has poor solubility in vitro, we resorted to use a more soluble N-terminal deletion of the protein, Pyd-PΔCC [36]. In salivary glands, both Pyd-PΔCC and Pyd-P colocalize with Scramb1-A-V5 in the plasma membrane (Fig. S4, B and C). Notably, Pyd-PΔCC was co-immunoprecipitated with Scramb1-A-V5, indicating an in vivo interaction between the two proteins (Fig. 4A). This interaction was further examined using a proximity labelling Bio-ID approach [37]. Pyd-P was fused to TurboID and expressed together with Scramb1-A-V5 in salivary glands. We found that Scramb1-A-V5 was biotinylated, indicating close proximity between the two proteins. In contrast, Scramb1-A-V5 was not biotinylated when coexpressed with TurboID-V5 as a control (Fig. 4B). These results support the hypothesis that Pyd helps recruit Scramb1-A to the SD pre-complexes. This is consistent with the observation that in nephrocytes of the null allele pydex147, Scramb1-A-V5 remains mostly cytoplasmic, with minimal overlap with Duf and Sns, which localize to cell-cell contact sites (Fig. 4C, arrows). In contrast, in dufsps1 nephrocytes, which lack SDs [10], Scramb1-A-V5 colocalizes with Pyd and Sns in cortical foci (Fig. S4 D).
Fig. 4
Scramb1-A interacts with Pyd. (A) Co-IP of Scramb1-A-V5 and Pyd-PΔCC from a lysate of salivary glands coexpressing both proteins. The same lysate was incubated with a magnetic matrix coupled to either anti-β-galactosidase in the control (ctrl) experiment or anti-V5 (V5) antibodies. The eluates were analyzed by western blot using anti-V5 to detect Scramb1-A-V5 and anti-Pyd. Pyd-PΔCC was notably elevated in the eluate from the V5 matrix compared to the control, which shows some unspecific Pyd binding to the matrix. (B) Proximity labeling with biotin of Scramb1-A-V5 by Pyd-P-TurboID-V5. Biotinylated proteins were isolated from lysates of salivary glands expressing Scramb1-A-V5 alongside Pyd-P-TurboID-V5 or TurboID-V5 (control). The lysates (input, 10% loaded) and purified fractions (P) were analyzed by western blot using anti-V5 antibody to detect TurboID-V5, Pyd-P-TurboID-V5 and Scramb1-A-V5. Scramb1-A-V5 was biotinylated by Pyd-P-TurboID-V5 but not by the control TurboID-V5, indicating a close association between Scramb1-A and Pyd. Notice that Pyd-P-TurboID-V5 and TurboID-V5 auto-biotinylate themselves. (C) Immunostaining of pydex147 nephrocytes expressing UAS-scramb1-A-V5 driven by pros-Gal4 to detect Scramb1-A-V5 (anti-V5 antibody), Sns and Duf, as indicated. Nuclei were labeled with DAPI (blue). pydex147 nephrocytes lack SDs and both Sns and Duf accumulate in regions of contact between aggregated nephrocytes (arrows) whereas Scramb1-A-V5 shows a cytoplasmic distribution
As mentioned above, isoform Scramb1-A, but not Scramb1-B, contains an N-terminal region of 183 amino acids that is rich in proline residues and that could potentially mediate its targeting to the SDs by interacting with Pyd. To investigate the significance of this region, we generated a UAS-scramb1-B-V5 transgene. Surprisingly, its expression in nephrocytes resulted in no detectable protein (Fig. 5, A and A’). However, it can readily be detected in wing imaginal discs after induction using the hh-Gal4 driver, even though at lower levels than Scramb1-A-V5 (Fig. S4, E and F). These results suggest that the UAS-scramb1-B-V5 transgene is translated, but the isoform Scramb1-B, lacking the proline-rich region, is unstable and degraded, particularly in nephrocytes. To test this possibility, we blocked both, the lysosomal and proteasome degradation pathways. Inhibiting lysosomal degradation by depleting the protein Vps18, which is involved in trafficking cargo to the late endosomes (dor8 mutant), led to the accumulation of Scramb1-B-V5 in cytoplasmic vesicles (Fig. 5, B and B’, arrows). Silencing several proteasome subunits by RNA interference, namely Prosα1, Prosα6 and Prosβ3 [38], resulted in a strong accumulation of Scramb1-B-V5 in cytoplasmic aggresomes, identified by anti-ubiquitin immunoreactivity (Fig. 5, C-D’, arrows; and Fig. S4 G). Thus, Scramb1-B is degraded in nephrocytes by the lysosomal and proteasomal pathways. Significantly, even after blocking its degradation, we did not observe colocalization with SD markers (Fig. 5, A-C’, arrowheads). This suggests that the proline-rich region of Scramb1-A is crucial for its stability and accumulation in SDs.
Fig. 5
Scramb1-A proline-rich domain is required for protein stability and localization to SDs. (A-C’) Nephrocytes overexpressing the isoform Scramb1-B-V5 driven by sns-GCN-Gal4 in an otherwise wild-type background (+), and in nephrocytes with compromised protein degradation via the lysosome pathway (dor8 mutants) or the proteasome pathway (Prosα1 silencing), immunostained to reveal Scramb1-B-V5 (anti-V5 antibody) and Pyd in medial sections, as indicated. Scramb1-B-V5 is undetectable in A (+), but accumulates in cytoplasmic vesicles in dor8 (arrows in B-B’) and in aggresomes in Prosα1 depleted nephrocytes (arrows in C-C’). Scramb1-B-V5 does not accumulate in SDs (arrowheads). (D-D’) Silencing the proteasome subunit Prosα6 in nephrocytes expressing UAS-scramb1-B-V5 (sns-GCN-Gal4) results in the formation of aggresomes, identified by the accumulation of Ubiquitin (arrows), that also contain Scramb1-B-V5. Medial sections are shown. (E-H’’) Nephrocytes expressing Scramb2-HA or the chimera Spro-scramb2-V5 (Scramb1-A proline-rich region fused to Scramb2), driven by sns-GCN-Gal4, stained with anti-Pyd and anti-HA or anti-V5, as indicated. E-E’’ and G-G’’ depict medial sections. The boxed regions in E and G are magnified in E’-E’’ and G’-G’’ respectively. Scramb2-HA accumulates at similar levels in the plasma membrane (arrows in E’-E’’) and the subcortical region, whereas Spro-scramb2-V5 accumulates at higher levels in the plasma membrane, colocalizing with Pyd (arrows in G’-G’’). The corresponding intensity profiles for Pyd and Scramb2-HA or Spro-scramb2-V5, expressed in arbitrary units, are shown in I and J, as indicated. The plasma membrane was registered at the 1 µm position. (F-F’’) Cortical section displaying partial colocalization between Pyd and Scramb2-HA, indicated by a Pearson’s colocalization coefficient of 0.335. (H-H’’) Cortical sections showing the distribution of Pyd and Spro-scramb2-V5’, colocalizing in a fingerprint-like pattern (arrows) with a Pearson’s colocalization coefficient of 0.516. Nuclei are labeled with DAPI (blue). A-C’ shown at the same magnification
Next, we tested whether the proline-rich region of Scramb1-A could confer targeting to SDs when fused to Drosophila Scramb2 in the chimera UAS-spro-scramb2-V5. Upon expression of UAS-scramb2-HA in nephrocytes, the protein accumulated to comparable levels in the subcortical region, where the labyrinthine channels are located, and in the plasma membrane (Fig. 5, E-E’’, arrows points to the plasma membrane; and I). In cortical sections, Scramb2-HA does not describe the fingerprint-like pattern characteristic of SDs (Fig. 5, F-F’’). In contrast, the chimeric protein displayed an enhanced signal in the plasma membrane, where it colocalized with Pyd (Fig. 5, G-G’’, arrows; and J). In cortical sections, Spro-scramb2-V5 colocalized with Pyd in a fingerprint-like pattern (Fig. 5, H-H’’, arrows). These findings provide additional support to the notion that the proline-rich region of Scramb1 promotes SD targeting. However, it is noteworthy that the expression of this chimeric protein in scramb143 nephrocytes failed to rescue its phenotype (Fig. S4, H and I).
The putative Ca2+ binding domain in Scramb1-A is required for SD assembly but not for its localization to SDsWe have shown that Scramb1-A localization to pre-SD complexes is required for SD assembly and, in particular, for Duf recruitment. However, the molecular role that Scramb1-A plays within the SD complex remains unclear. Homologous proteins, such as human PLSCR1, have been shown to have phospholipid scrambling activity in vitro and to participate in membrane-driven processes in several cellular contexts [31, 32, 39, 59]. These activities are regulated by Ca2+ binding to a short 12-residue sequence with homology to the loop region of EF hand domains [40], a region conserved in Scramb1 (Fig. S1 F; and Fig. S5 A). To examine the role of this putative Ca2+ binding domain, we engineered three transgenes, each containing a single mutation in conserved residues previously shown to be required for PLSCR1 activity [40]. Expression of Scramb1-AD372A-V5 or Scramb1-AF374A-V5 in scramb143 nephrocytes rescued SD formation at a similar level than the wild-type protein, as seen by the distribution of Duf in a characteristic fingerprint pattern in the cortical region and the absence of nephrocytes agglutination (Fig. S5, B-C’). The expression of Scramb1-AF378A-V5 resulted in a partial rescue of SD formation, with nephrocytes showing reduced SD density and cortical regions devoid of SDs (Fig. 6, A and B, white arrow). Interestingly, this mutant protein also accumulated in cortical foci that lacked or displayed low levels of Duf (Fig. 6B, green arrow).
Fig. 6
Requirement of the putative Ca2+-binding region of Scramb1-A. (A-B) Immunostaining of scramb143 nephrocytes partially rescued by the expression of Scramb1-AF378A-V5, containing one residue substitution within its putative Ca2+ binding region, driven by pros-Gal4 and shown at medial (A) and cortical (B) planes. The nephrocyte surface is partially covered by short SD strands identified by Duf accumulation, that coexist with foci containing Scramb1-AF378A-V5 and low levels of Duf (green arrow). White arrow in A points to a region devoid of SDs. (C-D4) Immunostaining of scramb143 nephrocytes expressing Scramb1-AD372A, F378A-V5, a variant containing two residue substitutions within its putative Ca2+ binding region, driven by pros-Gal4, shown at a medial (C) and a cortical view at a higher magnification (D). The highlighted region in D is also shown as single channels (D1-D4), as indicated. Very few SDs are formed. Pyd, Sns and Scramb1-AD372A, F378A-V5 (anti-V5 antibody) accumulate in abundant cortical foci that contain low levels of Duf. (E-E’’’) Cortical view of a first instar larval nephrocyte expressing Scramb1-AD372A, F378A-V5 (pros-Gal4), immunostained as indicated. Similarly to Scramb1-A, this Ca2+-insensitive variant accumulates in SDs, identified by Duf and Pyd co-expression. (A, C) Nuclei were labeled with DAPI (blue). D-D4 shown at the same magnification
Lastly, expression of Scramb1-AD372A, F378A-V5, a variant with mutations in two of the conserved residues, failed to rescue SD formation. Nephrocytes were agglutinated and lacked or displayed only a few SD strands. Moreover, a significant portion of the nephrocyte cortex was covered by foci that accumulated high levels of Scramb1-AD372A, F378A-V5, Pyd and Sns, but showed substantially reduced levels of Duf (Fig. 6, C-D4). These foci are similar to the ones observed in scramb143 mutant nephrocytes (compare with Fig. 3A). Together, these data indicate that Scramb1 putative Ca2+ binding region is required to promote SD formation but not for Scramb1 colocalization with SD components. In fact, when Scramb1-AD372A, F378A-V5 is expressed in a wild-type background, it partially accumulates in SDs (Fig. 6E-E’’’).
Palmitoylation of Scramb1 is essential to promote SD formationTo explore the cellular processes dependent on Scramb1 activity during SD formation, we undertook a proteomic approach to identify Scramb1 interactors. We generated a transgene expressing Scramb1-A fused with protein-A, enabling efficient purification of protein complexes by affinity chromatography. This fusion protein retains functionality, as it rescues SD formation in scramb143 nephrocytes (Fig. S5, D and E). We expressed Scramb1-A-protA in the larval fat body, a large organ of mesodermal origin, and purified it from larval extracts, identifying 72 co-purifying proteins not present in a control experiment using an empty matrix (Supplementary Table 1). 23 putative interactors were mitochondrial proteins, likely reflecting a mitochondrial role for scramb1 in the fat body, where it is also expressed. These interactors are unrelated to SD formation and therefore, were not further examined. We focused our attention on the interactors located at the plasma membrane and within the endo-lysosomal vesicular system, as they could potentially function alongside Scramb1 in promoting the assembly of SDs.
Among the potential Scramb1-A interactors at the plasma membrane was Flotillin2 (Flo2), a conserved protein found in cholesterol-rich membrane microdomains [41]. We validated this interaction through co-immunoprecipitation assays in salivary glands coexpressing UAS-scramb1-A-V5 and UAS-Flo2-RFP (Fig. 7C).
Fig. 7
Requirement of Scramb1-A palmitoylation sites. (A-B) Scheme of Scramb1-A domain composition, highlighting a cluster of conserved cysteine residues (yellow) matching the human PLSCR1 palmitoylation site and sequence alignment of the region (B). The construct UAS-Scramb1-ANP3-V5A contains mutations in residues 184, 188 and 189, in red. A conserved putative Ca2+- binding site (green) is also indicated. (C) Co-IP from salivary glands coexpressing Scramb1A-V5 and Flo2-RFP. The extract was incubated with a magnetic matrix coupled to anti-V5 or to anti-?-galactosidase as a control, and the eluates analyzed by western blot to detect Scramb1-A-V5 (anti-V5 antibody) and Flo2-RFP (anti-RFP), as indicated. Flo2-RFP was co-immunoprecipitated with Scramb1-V5. (D) Genetic interaction between scramb1 and Flo2. Three genotypes were quantitated: scramb1/scramb1 (n = 105 cells), scramb1/scramb1 (n = 87 cells) and a double mutant combination flo2/ Y; scramb1/scramb1 (n = 193 cells). Nephrocytes were immunostained for Duf and Pyd and classified into four categories, from no SD strands observed (0) to SDs covering the entire nephrocyte surface (3). Examples are shown in Fig. S7. A mutation in Flo2 normalizes the scramb1 phenotype. Asterisks show statistical significance (* P < 0.05, *** P < 0.001). (E-F’’) Subcellular localization of non-palmitoylable Scramb1-ANP3-V5 driven by pros-Gal4. Immunostainings with anti-V5 and anti-Pyd are shown in medial planes (E-E’) and in cortical planes at higher magnification (F-F’). Scramb1-ANP3-V5 colocalizes with Pyd in SDs (F-F’’) and also accumulates in nuclei, colocalizing with DAPI in blue (arrows). (G-G) scramb1 nephrocytes expressing UAS-scramb1-ANP3-V5 (pros-Gal4), immunostained to show the cortical distribution of Scramb1-ANP3-V5 (anti-V5 antibody), Sns, Duf and Pyd in foci and occasional short SD strands (yellow arrows). E-E’, F-F’’ and G-G shown at the same magnification
Supporting the interaction of Flo2 with Scramb1 in nephrocytes, overexpressed Flo2-RFP, in addition to a cytoplasmic distribution, colocalized with the SD marker Duf in the plasma membrane (Fig.
Comments (0)