
Remember me
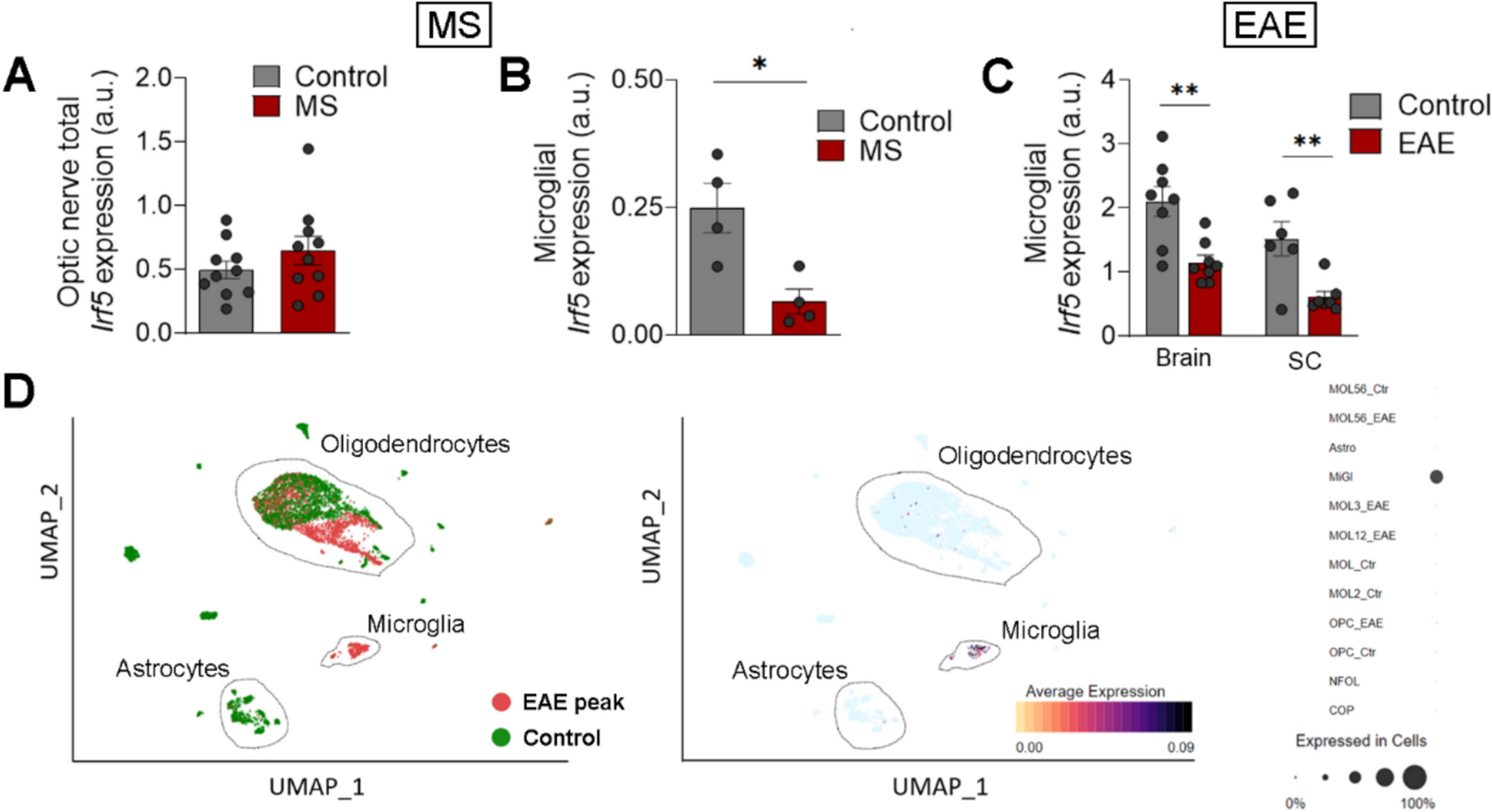


To explore the role of Smad3 in the cytokine and chemokine productions by NK cells, we firstly performed cytokine profiling with supernatant from empty vector (EV)-transfected NK-92 (NK-92-EV) cells and Smad3-silencing NK-92 (NK-92-S3KD) cells generated as described previously [16]. As shown in Fig. 1A, GM-CSF was the most remarkably changed cytokine among the 80 proteins in NK-92-S3KD, ten times higher than that in NK-92-EV supernatant. This indicated Smad3 may be a critical regulator for GM-CSF production in NK-92 cells. As Smad3 is one of the key transcription factors in canonical TGF-β signaling, we then evaluated the impact of TGF-β1 stimulation on the production of GM-CSF in NK-92 cells. Consistent with cytokine array results, silencing Smad3 substantially increased the level of GM-CSF produced by NK-92 cells (Fig. 1B-C). However, TGF-β1 stimulation inhibited the production of GM-CSF in both NK-92-EV and NK-92-S3KD (Fig. 1C), implying the participation of TGF-β-mediated Smad3-independent regulations.
Promoter analysis identified a predicted Smad binding site on the promoter region of CSF2 (GM-CSF) conserved between mouse and human with Evolutionary Conserved Regions (ECR) browser (rVista 2.0, https://rvista.dcode.org/) and JASPAR database (http://jaspar.genereg.net) (Fig. 1D) [32, 33]. Chromatin immunoprecipitation (ChIP) assay was performed thereupon to verify this transcription regulation. As shown in Fig. 1E, TGF-β1 stimulation enhanced the binding of p-Smad3 to the predicted Smad binding site on the promoter of CSF2. Dual luciferase reporter assay was subsequently carried out to validate whether Smad3 regulates the transcription of CSF2 through binding to the specific binding site. The transfection of a mutant Smad3 containing plasmid markedly promoted the transcription activity of the CSF2 promoter compared with the transfection of a Smad3 containing plasmid, indicating the negative regulation of GM-CSF expression by Smad3. Meanwhile, the introduction of a point mutation on the predicted Smad binding site on the CSF2 promoter also abrogated the suppression of Smad3 on the transcription activity of the CSF2 promoter (Fig. 1F). Taken together, Smad3 acted as a transcriptional repressor in TGF-β1-mediated suppression on GM-CSF production.
Fig. 1
TGF-β suppresses GM-CSF production in NK cells through Smad3-mediated transcriptional regulation. (A) Cytokine profiling detecting 80 different cytokines or chemokines in NK-92-EV and NK-92-S3KD supernatant. (B) GM-CSF levels in the supernatant of NK-92-EV and NK-92-S3KD cells determined by ELISA. Each bar represents the mean ± SD for groups of three independent experiments. *** p < 0.001 compared to NK-92-EV. (C) ELISA determining GM-CSF levels in NK-92-EV and NK-92-S3KD cells stimulated with recombinant TGF-β1 for 24 h. Each bar represents the mean ± SD for groups of three independent experiments. ** p < 0.01, *** p < 0.001 compared to no TGF-β stimulation; ### p < 0.001 as indicated. (D) Schematics of predicted Smad binding site on CSF2 gene. (E) ChIP assay verified the physical interaction between p-Smad3 and its predicted binding site in the promoter region of CSF2 gene. (F) Dual luciferase reporter assay validated Smad3 functions as a transcriptional repressor for CSF2 gene. wt-Smad3: pcDNA3.1 vector containing Smad3; mut-Smad3: pcDNA3.1 vector containing mutant Smad3. pGL3-basic: original pGL3 luciferase reporter basic vector; pGL3-CSF2: pGL3 vector containing CSF2 promoter sequence; pGL3-mut-CSF2: pGL3 vector containing CSF2 promoter sequence with a point mutation on the predicted Smad binding site. Each bar represents the mean ± SD for groups of three independent experiments. *** p < 0.001 compared to wt-Smad3
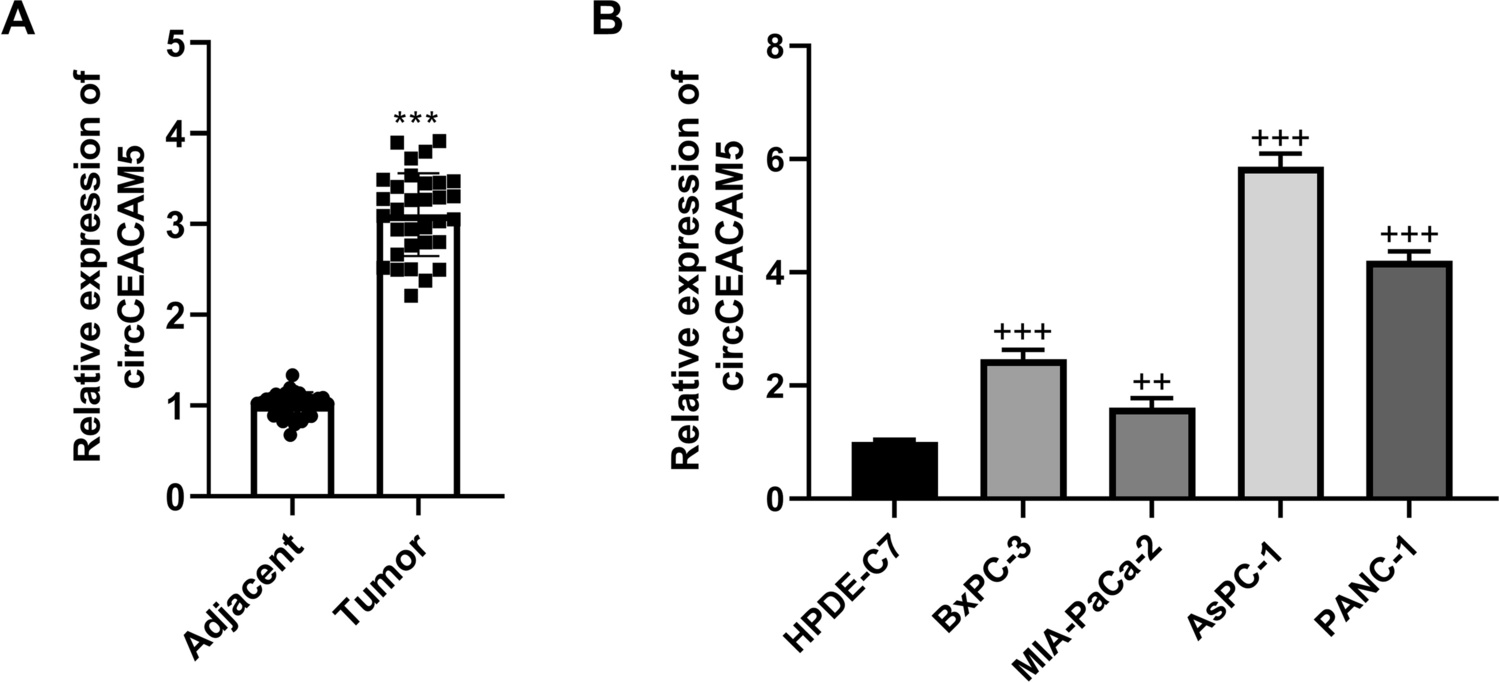
NK-derived GM-CSF suppresses lung carcinoma progressionGiven the dichotomic functions of GM-CSF in cancer progression, we then examined the role of NK-derived GM-CSF in the progression of syngeneic Lewis Lung Cancer (LLC) in C57BL/6 mice. LLC-bearing mice were treated with either BM-NK from Smad3 wild-type mice transfected with scramble sequence, BM-NK from Smad3 knockout mice transfected with scramble sequence or BM-NK from Smad3 knockout mice transfected with short interfering RNA (siRNA) targeting GM-CSF (si-GM-CSF) once a week. Neither deletion of Smad3 nor knockdown of GM-CSF significantly influenced NK cell proliferation or apoptosis, as determined by Annexin V-PI staining and MTT assay (Suppl. Figure 1 A-C). Consistent with our previous findings [16], tumor metabolic activity in mice receiving Smad3 knockout NK cell therapy was 80% lower than mice receiving Smad3 wild-type NK cell therapy as determined by bioluminescence imaging, suggesting Smad3 significantly suppresses the anti-cancer activity of NK cells. While the bioluminescence intensity in LLC-bearing mice receiving NK cell therapy was substantially promoted when GM-CSF was silenced in Smad3 knockout NK cells, rebounding to 72% of the intensity in mice receiving Smad3 wild-type NK cell therapy (Fig. 2A and B). Correspondingly, treatment with BM-NK from Smad3 knockout mice resulted in a 2.5-fold increase in the inhibition of LLC progression, which was voided by disrupting GM-CSF production as indicated by both tumor volume and tumor weight in Fig. 2C and D.
Fig. 2
Silencing GM-CSF diminishes the therapy effects of Smad3 knockout NK cells in mouse syngeneic LLC model. (A) Bioluminescence imaging and (B) corresponding quantitative analysis of luminescent intensity. (C) Tumor volume measured every 5 days. (D) Tumor weight at day 30 since the initiation of NK cell therapies. Each bar represents the mean ± SD for groups of eight mice. *** p < 0.001 compared to Ctrl; ## p < 0.01, ### p < 0.001 as indicated. S3 WT: Smad3 wild-type; S3 KO: Smad3 knockout; sc or scramble: scramble sequence; si-GM-CSF: siRNA targeting GM-CSF.
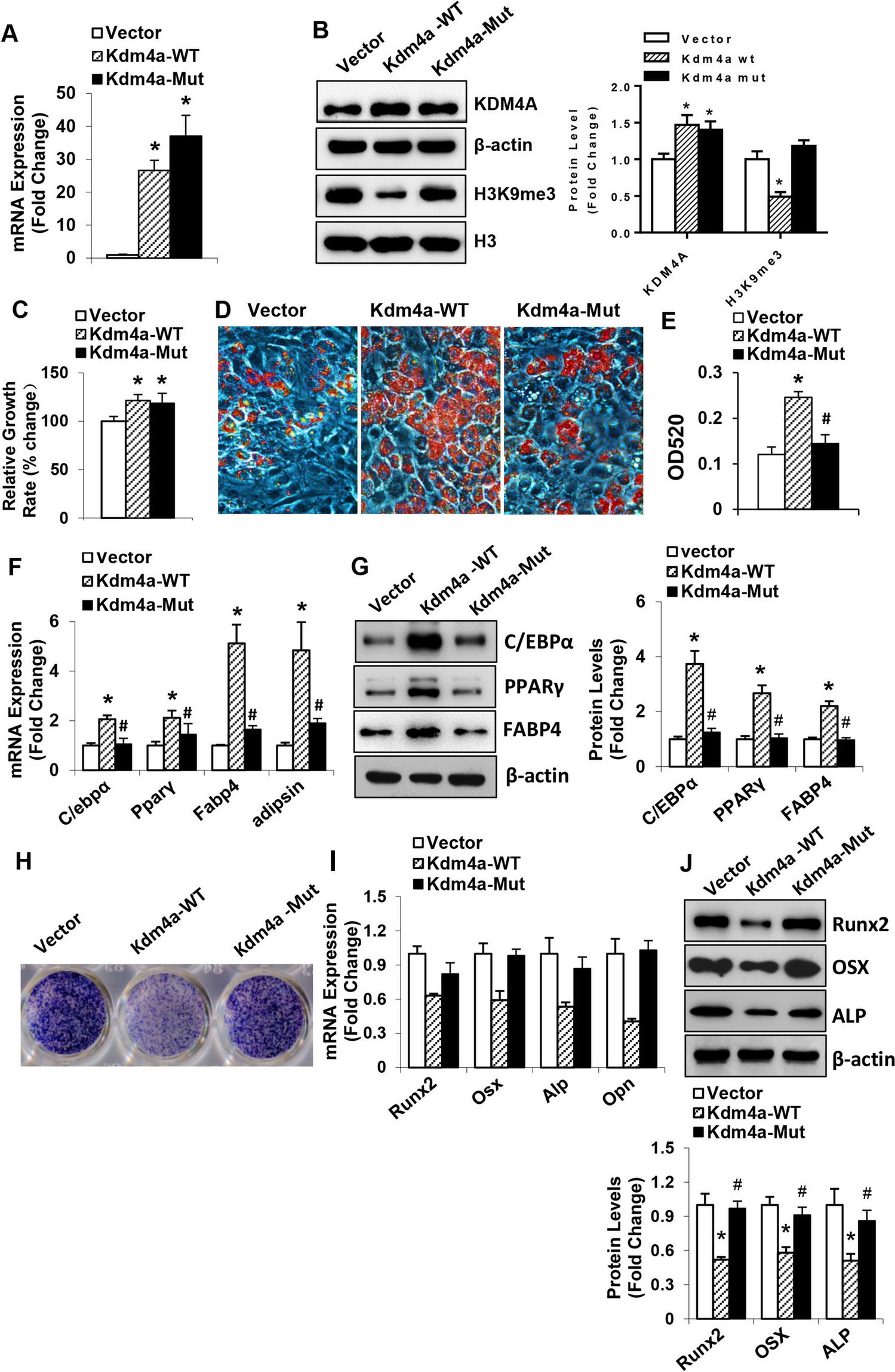
As shown in Fig. 3A, the production of GM-CSF by circulating NK cells was doubled in mice receiving Smad3 knockout-NK cell therapy compared with Smad3 wild-type NK cell therapy, however it was reduced in those receiving Smad3 knockout NK cell transfected with GM-CSF targeting siRNA. Whereas the proportions of GMCSF+ NK1.1− cells were not significantly changed in mice with or without NK cell therapies. In parallel, comparing with mice treated with Smad3 wild-type NK cells, the proportion of GM-CSF producing NK in the tumor microenvironment was largely promoted in those treated with Smad3 knockout NK cells, but reversed in mice treated with GM-CSF-silenced Smad3 knockout NK (Fig. 3B). It should be pointed out that NK cells detected by NK1.1 staining may include a tiny amount of NKT cells. However, as shown in Suppl. Figure 2A, the proportions of NKT cells within the BM-NK cells used for cell therapy were less than 2% across all three groups. Besides, very few NKT cells were found in the tumor microenvironment of mice receiving NK cell therapies (Suppl. Figure 2B).
Meanwhile, both tumoral and serum levels of GM-CSF in mice receiving Smad3 knockout NK therapy were much higher than that in mice receiving Smad3 wild-type NK therapy, whereas they were considerably reduced in mice receiving GM-CSF-silenced Smad3 knockout NK therapy (Fig. 3C and D), illustrating NK cell as an important source for GM-CSF in LLC-bearing mice. Altogether, silencing GM-CSF in Smad3 knockout NK cells remarkably mitigated the therapeutic effects of Smad3 knockout NK cell therapy, highlighting the significance of GM-CSF in NK-mediated anti-cancer effects.
Fig. 3
The level of GM-CSF is substantially decreased in mice receiving GM-CSF-silenced Smad3 knockout NK cell therapy. (A) Flow cytometry detecting GM-CSF-producing NK cells in the circulation of LLC-bearing mice receiving NK cell therapies, and corresponding quantifications. (B) Immunofluorescence staining detecting GM-CSF-producing NK in LLC tumor microenvironment and corresponding quantification. Scale bar, 50 μm. The levels of GM-CSF in (C) tumor tissue and (D) serum in LLC-bearing mice receiving NK cell therapies determined by ELISA. Each bar represents the mean ± SD for groups of five to six mice. * p < 0.05, ** p < 0.01, *** p < 0.001 compared to Ctrl; ## p < 0.01, ### p < 0.001 as indicated. S3 WT: Smad3 wild-type; S3 KO: Smad3 knockout; sc or scramble: scramble sequence; si-GM-CSF: siRNA targeting GM-CSF.
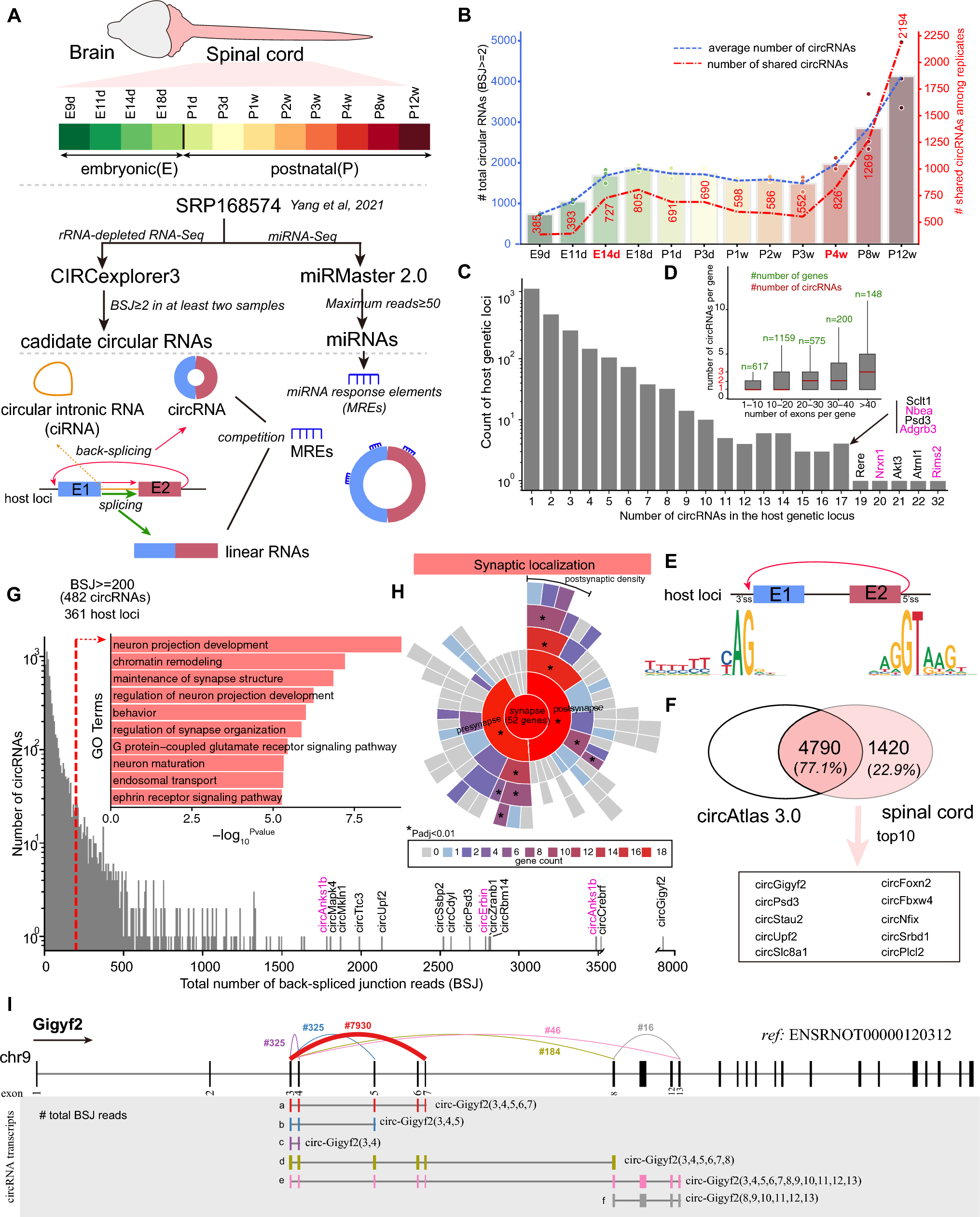
NK-derived GM-CSF promotes DC- and M1-mediated recruitment and activation of effector T cellsAs GM-CSFR is mostly expressed on myeloid cells such as DC, monocytes and macrophages, the anti-cancer GM-CSF effects are largely attributed to the influx and activation of DC and pro-inflammatory M1 macrophages, which subsequently activate T cells through antigen presenting in the tumor tissue [34]. Therefore, we evaluated the impact of silencing GM-CSF in Smad3 knockout NK cells as a cell therapy on the recruitment of DC, M1 macrophages and T cells in LLC microenvironment. In comparison with Smad3 wild-type NK cell therapy, the accumulation of DC in LLC tumor was increased twofold by Smad3 knockout NK cell therapy (Fig. 4A). Nevertheless, this elevation was abolished by the knockdown of GM-CSF in Smad3 knockout NK cells. Similarly, the number of tumor-infiltrated iNOS+ M1 was markedly increased in mice receiving Smad3 knockout NK cell therapy as compared with those receiving Smad3 wild-type NK cell therapy. Likewise, reducing NK-derived GM-CSF with si-RNA also nullified the protective effects of Smad3 knockout NK cell therapy on the recruitment of M1 macrophages, which was aligned with the changes of GM-CSF levels in the tumor tissue (Fig. 3C).
Compared with mice treated with Smad3 wild-type NK cells, the recruitment of CD8+ T cells was largely enhanced in mice treated with Smad3 knockout NK cells. Moreover, the proportion of effector CD8+ T cells was additionally influenced by NK-derived GM-CSF, as silencing GM-CSF in Smad3 knockout NK cells notably abrogated the recruitment of CD8+ T cells (Fig. 4C). Overall, Smad3-mediated inhibition of GM-CSF production in NK cells significantly impeded the recruitment of DCs and M1 macrophages in LLC tumor, which consequently reduced effector T cells in LLC microenvironment.
Fig. 4
NK-derived GM-CSF encourages DC- and M1 macrophage-mediated CD8+ T cell activation and recruitment in lung carcinoma. Immunofluorescence staining detecting (A) the accumulation of DC in LLC tumor; (B) the accumulation of M1 macrophages in LLC tumor; and (C) the accumulation of CD8+ effector T cells in LLC tumor. Scale bar, 50 μm. Each bar represents the mean ± SD for groups of five to six mice. ** p < 0.01, *** p < 0.001 compared to Ctrl; ## p < 0.01, ### p < 0.001 as indicated. S3 WT: Smad3 wild-type; S3 KO: Smad3 knockout; sc or scramble: scramble sequence; si-GM-CSF: siRNA targeting GM-CSF.
As high dose GM-CSF may play a pathogenic role by stimulating the differentiation of immunosuppressive MDSC and Treg cells [35], we further evaluated the influence of NK cell therapies on the accumulation of MDSCs and Tregs in the LLC tumor tissue. As shown in Suppl. Figure 3A, the amount of tumor-infiltrated MDSC was enhanced by Smad3 knockout NK cell therapy from 17 to 23%, but not by Smad3 wild-type NK cell therapy nor GM-CSF-silenced Smad3 knockout NK cell therapy. Meanwhile, the percentage of Treg cells was increased from 4.7 to 5.7% compared with control by Smad3 wild-type NK cell therapy, and to 7.2% by Smad3 knockout NK therapy, while blocking GM-CSF production by these NK cells largely mitigated the pathogenic effect, resulting in a reduction in Treg proportion to 5.5% of total tumor-infiltrated leukocytes (Suppl. Figure 3B).
NK-derived GM-CSF promotes the survival of tumor-infiltrated neutrophils, which subsequently triggers the terminal maturation and promotes the anti-cancer functions of NK cells in positive feedback circuitIn addition to T cells, NK cells also serve as potent tumor killers through direct killing and cytokine-mediated cellular crosstalk. To explore whether NK-derived GM-CSF also regulates NK-mediated immune responses in the tumor microenvironment, we analyzed the effector functions of NK cells in LLC-bearing mice after 3-week NK cell therapies. As shown in Fig. 5A, the proportion of IFN-γ producing NK in the circulation of mice treated with Smad3 knockout NK cells was doubled compared to those treated with Smad3 wild-type NK cells, yet the proportion was decreased to half when GM-CSF was knocked down in the Smad3 knockout NK cells. We also observed the proportion of IFN-γ+ NK1.1− cells was remarkably enhanced by Smad3 knockout NK cell therapy compared with wild-type NK therapy, but it was largely reduced by GM-CSF knockdown. As the numbers of NKT cells (Fig. Suppl. Figure 2), CD8− T cells (Fig. 4) and Treg cells (Fig. Suppl. Figure 3B) were not remarkably changed by NK cell therapies, it is possible that the majority of the IFN-γ+ NK1.1− cells in Fig. 5A are cytotoxic T cells. To confirm this, IFN-γ and CD8 were co-stained in the tumor microenvironment. As shown in Suppl. Figure 4, the amount of IFN-γ-producing CD8+ T cells in the tumor tissue remarkably increased in mice receiving Smad3 knockout NK cell therapy compared with that receiving Smad3 wild-type NK therapy, while the increase was reduced by knocking down GM-CSF in Smad3 knockout NK cells.
Comparably, the percentage of cytolytic NK cells among total NK cells, characterized by the productions of granzyme B and perforin, in mice receiving GM-CSF-silenced Smad3 knockout NK cells, was also notably reduced to 33% of those receiving scramble sequence-transfected Smad3 knockout NK cells (Fig. 5B). Since the amount of effector T cells and cytolytic NK cells, being the two major sources of IFN-γ, granzyme B and perforin, were correlated with the production of GM-CSF by NK cells used for adoptive transfer therapies, we hypothesized that the levels of these cytokines would also be associated with the level of NK-derived GM-CSF in the tumor microenvironment. As shown in Fig. 5C-E, the enhancive effects on the production of IFN-γ, granzyme B and perforin induced by disrupting Smad3 in NK cells were largely impaired by knocking down GM-CSF, implying that NK-derived GM-CSF was crucial for NK- and cytotoxic T cell-mediated cytotoxicity in LLC microenvironment.
Fig. 5
GM-CSF augments the cytotoxic functions of NK cells in LLC-bearing mice. (A) Flow cytometry examining the proportion of IFN-γ producing NK in the circulation. (B) Flow cytometry detecting the proportion of Granzyme B and perforin producing cytolytic NK in the circulation (C-E) The levels of (C) IFN-γ, (D) granzyme B and (E) perforin in LLC tumor were analyzed by ELISA. Each bar represents the mean ± SD for groups of five to six mice. * p < 0.05, ** p < 0.01, *** p < 0.001 compared to Ctrl; # p < 0.05, ## p < 0.01, ### p < 0.001 as indicated. S3 WT: Smad3 wild-type; S3 KO: Smad3 knockout; sc or scramble: scramble sequence; si-GM-CSF: siRNA targeting GM-CSF.
To determine whether GM-CSF produced by NK cells can in turn directly regulate the cytotoxic functions of NK cells, the productions of IFN-γ, granzyme B and perforin in GM-CSF-silenced BM-NK cells were assessed by ELISA. According to the results in Suppl. Figure 5A-C, the levels of IFN-γ, granzyme B and perforin in the supernatant of both GM-CSF-silenced (si-GM) Smad3 wild-type and Smad3 knockout BM-NK cells were comparable to those in BM-NK cells transfected with scramble sequence. Moreover, stimulation with mouse recombinant GM-CSF also did not affect the production of these cytotoxic effectors by either Smad3 wild-type or Smad3 knockout BM-NK cells (Suppl. Figure 5D-F). These results suggested that GM-CSF could not regulate the effector functions of NK cells directly.
Jaeger et al. revealed that neutrophils play a critical role in the terminal maturation of NK cells, characterized by the expression of CD11b and CD43, and their corresponding effectiveness against cancer [36]. Through the interaction of CD18/ICAM-3, neutrophils directly bind to NK cells, accelerating their terminal maturation and promoting IFN-γ production [37]. Meanwhile, GM-CSF promotes the survival of neutrophil by suppressing their apoptosis [38]. These lead us to the hypothesis that NK-derived GM-CSF prolongs the lifespan of neutrophils, which consequently facilitates the maturation and boosts the cytotoxicity of NK cell to form a positive anti-cancer feedback loop. To validate this hypothesis, we determined the amount of CD18+ neutrophils in the circulation and tumor tissue after NK cell therapies. Compared with mice without treatment, cell therapy with Smad3 wild-type NK cells increased the number of neutrophils in circulation from 5 to 12%, and the amount was further doubled by treatment with Smad3 knockout NK cells (Fig. 6A). However, silencing GM-CSF in Smad3 knockout NK cells significantly blunted the protective effect of these NK cells, which lowered the proportion of CD18+ neutrophils from 25% back to 15.6%, indicating NK-derived GM-CSF was important for CD18+ neutrophil survival in the circulation. Similar effects were also observed in the LLC tumor, where the accumulation of CD18+ neutrophils was significantly enhanced by Smad3 knockout NK therapy compared with Smad3 wild-type NK therapy. While the number was substantially dropped when GM-CSF was silenced in Smad3 knockout NK therapy, which was even lower than that in mice treated with Smad3 wild-type NK therapy (Fig. 6B). Thereby, our data suggested NK cell prolonged the survival of CD18+ neutrophils in LLC-bearing mice by producing GM-CSF.
Fig. 6
NK-derived GM-CSF is essential for CD18+ neutrophil survival in LLC-bearing mice. (A) The proportion of CD18+ Ly6G+ neutrophils in the circulation analyzed by flow cytometry. (B) The proportion of tumor-infiltrated CD18+ Ly6G+ neutrophils in LLC-bearing mice analyzed by immunofluorescence. Each bar represents the mean ± SD for groups of five to six mice. ** p < 0.01, *** p < 0.001 compared to Ctrl; ## p < 0.01, ### p < 0.001 as indicated. S3 WT: Smad3 wild-type; S3 KO: Smad3 knockout; sc or scramble: scramble sequence; si-GM-CSF: siRNA targeting GM-CSF.
Next, we examined whether the terminal maturation of NK cells was affected by the level of NK-derived GM-CSF in the LLC mice. As shown in Fig. 7A, the proportion of CD11b+ CD43+ matured NK cells in total NK cells in the circulation of LLC-bearing mice was raised from 9 to 26% by Smad3 wild-type NK cell therapy, and it was further elevated to 48% by Smad3 knockout NK cell therapy. However, silencing GM-CSF in Smad3 knockout NK cells remarkably reduced the proportion of matured NK cells to only 20%, indicating that NK-derived GM-CSF was critical for the terminal maturation of circulating NK cells in LLC-bearing mice. Although the number of tumor-infiltrated NK 1.1+ cells was comparable in mice treated with either NK cell therapy, the percentages of matured CD11b+ CD43+ NK in both tumor-infiltrated NK cells and tumor-infiltrated CD45+ leukocytes were increased twofold by Smad3 knockout NK cell therapy compared with Smad3 wild-type NK cell therapy, while the increments were also abolished when GM-CSF was knocked down in Smad3 knockout NK cells (Fig. 7B). Such findings supported our hypothesis that NK-derived GM-CSF was essential for neutrophil survival, which subsequently encouraged the terminal maturation of NK cells and enhanced their effector functions against cancer.
Fig. 7
Silencing GM-CSF in Smad3 knockout NK cells used for cell therapy impedes the maturation of NK cells in LLC-bearing mice. (A) The proportion of CD11b+ CD43+ matured NK in the circulation and corresponding quantifications analyzed by flow cytometry. (B) The proportion of CD11b+ CD43+ matured NK in LLC tumor and corresponding quantifications analyzed by flow cytometry. Each bar represents the mean ± SD for groups of five to six mice. * p < 0.05, ** p < 0.01, *** p < 0.001 compared to Ctrl; # p < 0.05, ## p < 0.01, ### p < 0.001 as indicated. S3 WT: Smad3 wild-type; S3 KO: Smad3 knockout; sc or scramble: scramble sequence; si-GM-CSF: siRNA targeting GM-CSF.
Comments (0)