
Remember me

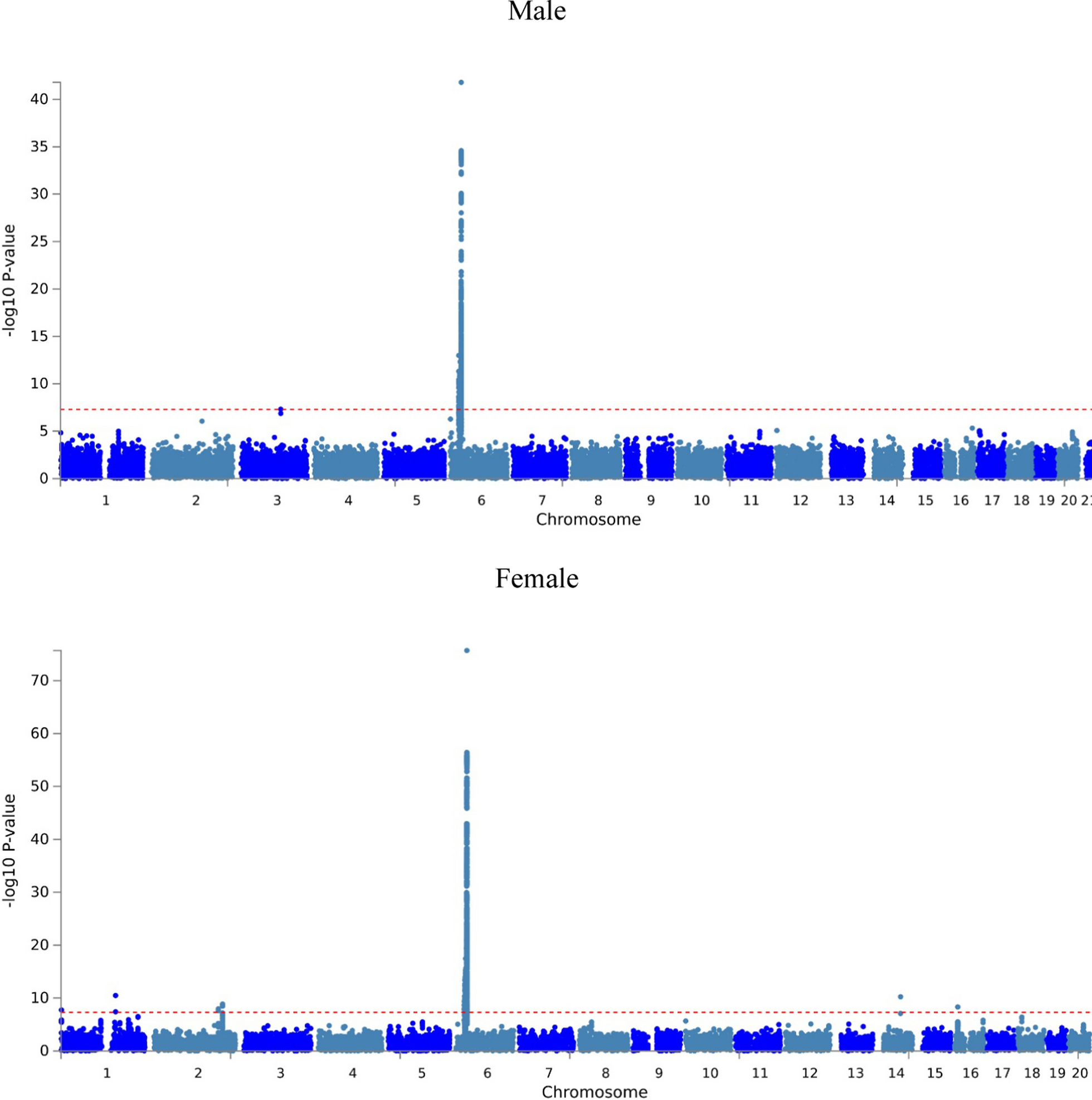



Consistent with previous studies [47, 48, 58], LB decreased body weight in P17 male and female pups (Additional file 1: Fig S2 A, B). Stereological analysis indicated that LB decreased the size of the granule cell layer (GCL) and the Cornu Ammonis (CA) pyramidal layer even after correction for body weight, with similar outcomes observed in males and females (Additional file 1: Fig S2C). The DNA, RNA, and protein contents extracted from the hippocampus were also reduced in male and female P17 LB pups (Additional file 1: Fig S2D). These findings replicate previous research demonstrating that LB reduces hippocampal volume in prepubescent pups [22, 47].
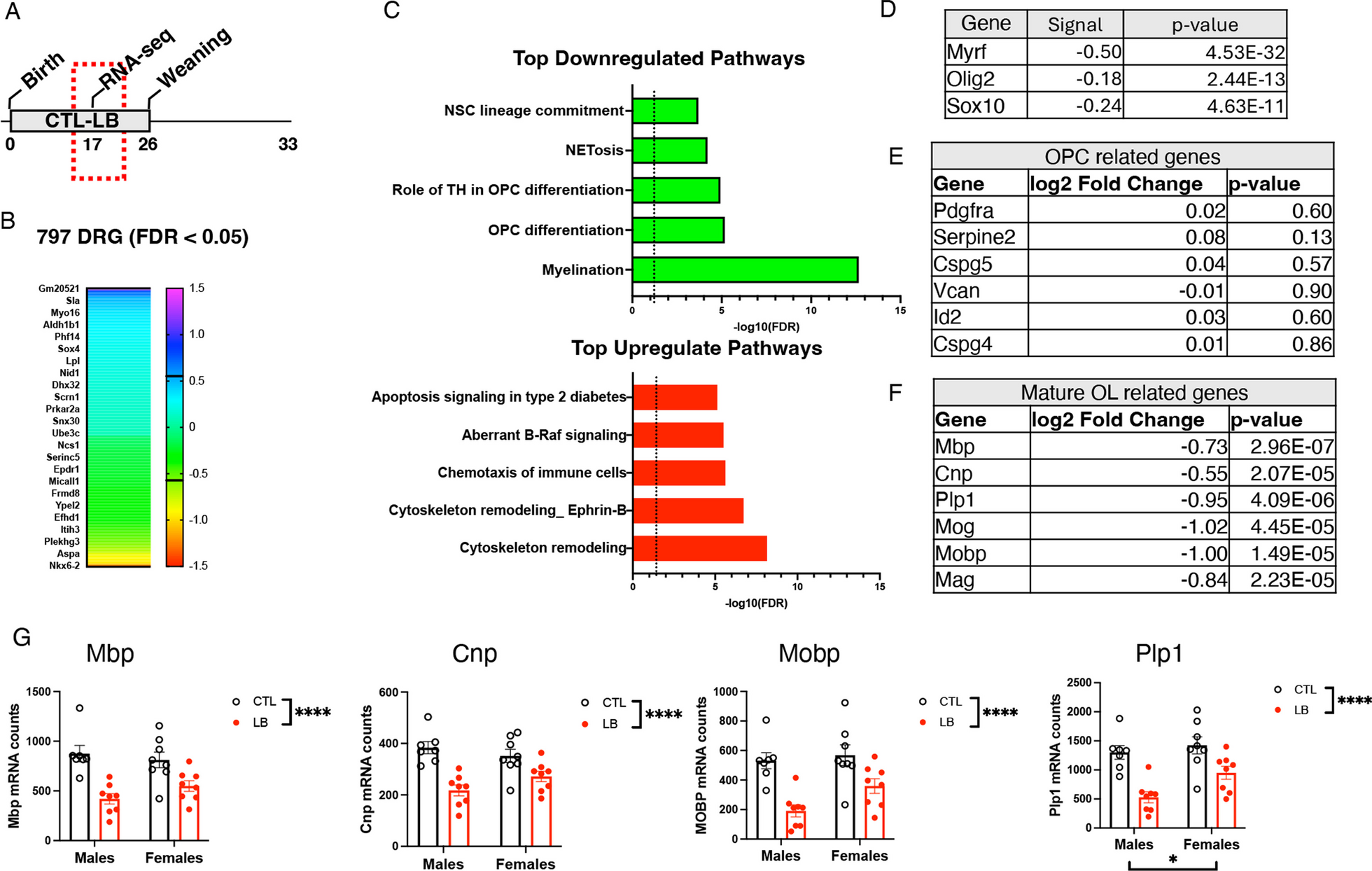
Bulk RNA-seq was used to further elucidate the mechanisms by which LB alters hippocampal development (Fig. 1A). Similar changes in gene expression were observed in male and female LB mice prompting us to focus on rearing-induced changes in gene expression. Using this approach, we identified 797 differentially regulated genes (DRGs) between CTL and LB (FDR < 0.05), most of which had a small effect size (i.e., log2 < 0.4, green/turquoise heatmap, Fig. 1B). Pathway analysis identified oligodendrocyte differentiation and myelination as the most significantly downregulated pathways in LB, whereas cytoskeleton remodeling was the most upregulated pathway in LB (Fig. 1C). The 20 most upregulated and downregulated genes are shown in Additional file 1: Fig S2. Transcription factor analysis identified reduced activity of Myrf, Olig2, and Sox10 as the key regulators of abnormal myelination in LB mice (Fig. 1D). Since Myrf is necessary for oligodendrocyte differentiation [53, 62], we assessed the effects of LB on genes that are specifically expressed in oligodendrocyte progenitor cells (OPCs) and mature oligodendrocytes (OLs). LB had no impact on OPC-specific genes (Fig. 1E) but significantly reduced the expression of OL-specific genes (Fig. 1F), indicating that LB impairs the differentiation of OPCs in the developing hippocampus. A 2 × 2 ANOVA examining the impact of rearing and sex on some of the most highly expressed OL-specific genes confirmed a highly significant effect of rearing (Fig. 1G). No significant effect of sex or interaction were found, except for Plp1 mRNA, where a significant effect of sex was observed. Nevertheless, a preplanned analyses of effect sizes found greater deficits in LB males (Fig. 1G. Cohen’s d effect sizes MBP: males = 2.45 vs females = 1.3, CNP: males = 2.71 vs females = 1.22, MOBP: males = 2.61 vs females = 1.21, Plp1: males = 2.66 vs females = 1.28).
Fig. 1
LB inhibits oligodendrocyte differentiation in the hippocampus of P17 prepubescent pups. A Experimental timeline. B Heatmap showing the effect size of differentially regulated genes (DRGs) at FDR < 0.05 between CTL and LB. C Pathway analysis of the top downregulated and upregulated pathways affected by LB. D Transcription factor analysis identified changes in Myrf, Olig2 and Sox10 activity as the most dysregulated factors affected by LB. LB does not affect the expression of OPC-specific genes E but significantly decreases the expression of genes expressed in mature oligodendrocytes (F–G). N = 7–8 mice per rearing and sex group. Error bars represent mean ± SEM. *p < 0.05, **p < 0.01, ****p < 0.0001
To further validate the RNA-seq data, we assessed the effects of LB on oligodendrocyte maturation in the stratum lacunosum moleculare (SLM), one of the most myelinated regions in the developing hippocampus (Fig. 2A) [27, 63, 64]. This was done by quantifying levels of MBP as a marker of myelination, number of PDGFRα-positive cells (OPCs), and number of CC1 positive cells (OL) in the SLM of P17 pups. Consistent with the RNA-seq data, we found a significant reduction in MBP staining in LB compared to control (F (1, 16) = 15.42, P = 0.0012, ηp2 = 0.49) with no significant effects of sex or interaction (Fig. 2B, C). There was also a significant reduction in the number of PDGFRα-positive cells in LB pups (Fig. 2B, D, rearing: F (1, 16) = 5.88, P = 0.0275, ηp2 = 0.27, sex: F (1, 16) = 0.05, P = 0.8, interaction: F (1, 16) = 0.01, P = 0.91). For CC1-positive cells (i.e., mature-OL), there was a significant effect of rearing (F (1, 16) = 35.56, P < 0.0001, ηp2 = 0.69) and sex (F (1, 16) = 4.664 P = 0.046). No significant interaction between rearing and sex was found but LB males showed a significantly greater reduction compared to LB female littermates (Cohen’s d effect size males = 4.58, females = 1.67, Fig. 2E). Since LB reduced both OPC and OL in the SLM, an OPC differentiation index was generated by dividing the number of CC1-positive cells by the number of PDGFRα-positive cells. There was a significant reduction in the OPC differentiation index in LB mice (F (1, 16) = 32.4, P < 0.000, ηp2 = 0.67, Fig. 2F) indicating that impaired OPC differentiation is primarily responsible for the reduced myelination in the SLM. Again, no significant effect of sex or interaction were found, but males were more severely impacted compared to females (Cohen’s d effect size males = 3.99, females = 1.50, Fig. 2F).
Fig. 2
OPC differentiation in the SLM is impaired in P17 Prepubescent LB mice. A Low (4X) and higher (20X) magnification of myelin basic protein (MBP) staining in the SLM of P17 pups. B Representative images of PDGFRα-positive OPCs, CC1-positive mature oligodendrocytes, and MBP in the SLM. Quantification of MBP (C), PDGFRα-positive OPCs (D), CC1 mature oligodendrocytes (E) and OPC differentiation index (F) in the SLM group. N = 5–6 mice per rearing and sex group. Error bars represent mean ± SEM. *p < 0.05, **p < 0.01, ****p < 0.0001
Axonal staining is reduced in the SLM of P17 LB miceTo test whether the reduced myelination is due to abnormal axonal innervation, we probed the hippocampus with anti-phosphorylated neurofilament H antibodies (NF-H) to quantify axonal fibers in the SLM of P17 control and LB mice (Fig. 3A). There was a significant reduction in NF-H staining in LB mice (F (1, 16) = 11.38, P = 0.0039), with no significant effect of sex and a trend for rearing by sex interaction (F (1, 16) = 3.30, P = 0.088). The trend in interaction was due to a greater reduction in NF-H staining in LB males compared to LB females (Fig. 3B, Cohen’s d effect size males = 2.05, females = 0.82). A similar pattern was observed for MBP staining, with reduced myelination in LB males (P < 0.0001, Cohen’s d = 3.23) but not in females (P = 0.48, Cohen’s d = 1.11, interaction: F (1, 16) = 14.20, P = 0.0017, Fig. 3C). The percentage of neurofilaments co-stained with MBP was reduced in LB compared to CTL (F (1, 16) = 5.78, P = 0.029), with no significant effects of sex or interaction, but a larger impact in males compared to females (Cohen’s d effect size males = 2.28, females = 0.45, Fig. 3D). These results suggest that reduced axonal availability contributes to the abnormal myelination observed in the SLM of 17-day-old prepubescent LB mice, with males being more impacted than females.
Fig. 3
LB causes a more pronounced reduction in axonal staining in the SLM of Prepubescent 17-day-old LB male mice. A Representative images of axonal staining using anti-NF-H and anti-MBP antibodies in the SLM. Quantification of NF-H staining (B), MBP staining (C) and NF-H-MBP colocalization (D). Scale bars in Fig. 4A are 50 microns. N = 5 mice per rearing and sex. Error bars represent the mean ± SEM. *p < 0.05, **p < 0.01, ****p < 0.0001
Since LB prepubescent males show more pronounced deficits in myelination, we used electron microscopy to further characterize myelination in the SLM of P17 LB males. This investigation confirmed reduced axonal myelination in the SLM of 17-day-old LB males (Fig. 4A). LB males exhibited a significant increase in the G-ratio (t (118) = 15.57, P < 0.0001, Cohen's d = 2.84, Fig. 4B). Axonal diameter was also increased in LB compared to CTL males (CTL = 1.05 ± 0.04 µm, LB = 1.47 ± 0.074 µm, t (95) = 4.87, P < 0.001, Cohen's d = 0.89), indicating that the reduced myelination was not due to reduced axonal diameter. Furthermore, the increase in the G-ratio observed in LB remained significant even after adjusting for the increase in axonal diameter (ANCOVA, F = 130.759, P < 0.001, Fig. 4C). This indicates that the elevated G-ratio seen in LB males is mainly influenced by decreased myelination.
Fig. 4
Transmission electron microscopy of myelinated axons in the SLM of P17 Prepubescent CTL and LB male mice. A Left, a light microscopy image of the same toluidine-stained sample to identify the SLM; Right, TEM images of the SLM in CTL and LB mice (visible axons indicated with red dots). B Image analysis of covariance between the G-ratio and axonal diameter (C). N = 20 axons per mouse with 3 male mice per condition. Error bars represent the mean ± SEM. ****p < 0.0001
Adolescent P29 LB mice are hyperactive and show extensive cortical atrophyWe have recently proposed that LB is a mouse model of childhood deprivation and neglect [1, 48]. Given that hyperactivity and extensive cortical thinning are consistent findings in children and adolescents exposed to severe social and cognitive deprivation [65, 66], we used the open field test and high-resolution dMRI to evaluate these two outcomes in P29 adolescent male and female mice (Fig. 5A). A 2 × 2 ANOVA for body weight revealed a significant effect of sex (F (1, 67) = 14.46, P = 0.0003), with males being larger than females (Fig. 5B), consistent with previous research indicating that puberty initiated at around this age in mice [39,40,41,42,43]. No significant effects of rearing or interaction were observed for body weight at this age (Fig. 5B). LB male and female P29 adolescent mice showed a significant increase in total distance traveled in the open field test (F (1, 65) = 3.95, P = 0.05), with no significant effect of sex or interaction (Fig. 5C). There was a significant interaction between rearing and sex for the time spent in the center (interaction: F (1, 65) = 7.71, P = 0.0072, Fig. 5D) and frequency of entering the center (interaction: F (1, 71) = 3.92, P = 0.05, Fig. 5E) due to an increased entry into the center in LB females but not LB males. These findings replicate our previous research in adolescent LB mice [47] and confirm increased hyperactivity in LB adolescent male and female mice.
Fig. 5
Adolescent P29 LB mice are hyperactive and show extensive reduction in cortical and subcortical gray matter. A Experimental timeline. Effects of rearing and sex on body weight (B), and exploratory behavior in the open filed test in P29 adolescent mice (C–E). CTL males N = 15, CTL females N = 14, LB males N = 16, LB females N = 24. F High resolution ex vivo dMRI found significant effects of rearing on volumetric changes using (minimal cluster size > 25 voxels, FDR < 0.1, p < 0.005). Areas with significant volume reduction are shown in red. N = 6 per rearing and sex. G Effects of rearing and sex on CA1 and dentate gyrus volumes. H–J The left entorhinal cortex was the only region showing a significant rearing by sex interaction (minimal cluster size > 5 voxels, FDR < 0.1, p < 0.0023). AI: Insular cortex, CB: Cerebellum, cpd: Cerebral peduncle, ENT: Entorhinal cortex, GP: Globus pallidus, HB: Hindbrain, HPF: Hippocampus, HY: HY: Hypothalamus, Int: Internal capsule, LSX: Lateral septal complex, MO: Motor cortex, OLF: Olfactory area, opt: Optic tract, OT: Olfactory tubercle, PFC: Prefrontal cortex, PTLp: Posterior parietal association cortex, RSP: Retrosplenial cortex, SC: Superior colliculus, SS: Somatosensory cortex, STR: Striatum, TH Thalamus, VIS: Visual cortex
Next, we conducted a 2 × 2 whole-brain voxel-based analysis to identify volumetric changes induced by rearing, sex, and their interaction [58, 59]. This unbiased approach revealed extensive volumetric reduction in LB adolescent male and female mice that included the sensory and motor cortex, prefrontal cortex, hippocampus, amygdala, and thalamus (shown in red in Fig. 5F). Follow-up region of interest analysis confirmed reduced CA1 (F (1, 20) = 15.25, P = 0.0009) and dentate gyrus volumes (F (1, 20) = 13.37, P = 0.0016) in both LB male and female P29 mice (Fig. 5G). These findings indicate that the lower hippocampal volume, seen in P17 LB pups (Additional file 1: Fig S2), persists in adolescent mice and is likely to represent global atrophy across multiple cortical and subcortical gray matter regions. Few brain regions showed a significant effect of sex (Additional file 1: Fig S4), and the only brain region that showed significant rearing by sex interaction was the left entorhinal cortex (Fig. 5H). A region of interest analysis confirmed a significant sex-by-rearing interaction in the left entorhinal cortex (F (1, 20) = 4.397, P = 0.049) driven by reduced volume in LB males (P = 0.0015, Cohen’s d = 2.18) but not LB females (P = 0.54, Cohen’s d = 0.61, Fig. 5I). A similar analysis in the right entorhinal cortex found a significant rearing effect (F (1, 20) = 11.85, P = 0.0026). No significant impact of sex (F (1, 20) = 0.61, P = 0.44) or interaction (F (1, 20) = 1.39, P = 0.25) were found, but the effect size in males was again larger (Cohen’s d effect size males = 1.72, females = 1.03, Fig. 5J).
Reduced axonal innervation persists in P33 adolescent mice and correlates with deficits in contextual fear conditioningNext, we tested whether the reduction in myelination and axonal staining seen in the SLM of P17 pups persisted and contributed to hippocampus-dependent deficits in adolescent mice. This was done by first testing contextual freezing in P31-33 adolescent mice and then assessing axonal staining and myelination in the SLM (Fig. 6A). Mice showed the expected increase in freezing in response to shock on the first day of training (F (2.958, 136.1) = 208, P < 0.0001). However, there was a significant interaction between rearing and sex for contextual freezing during the second day (F (1, 62) = 3.994, P = 0.05), driven by a significant reduction in freezing behavior in LB males (P = 0.0008, Cohen’s d = 1.55) but not LB females (P = 0.43, Cohen’s d = 0.36, Fig. 6B). Examination of NF-H staining in the SLM revealed significant effects of rearing (F (1, 20) = 9.55, P = 0.0058) and sex (F (1, 20) = 4.46, P = 0.047) but no significant interaction (F (1, 20) = 0.426, P = 0.52) (Fig. 6C). Nevertheless, as with the axonal staining at P17 (Fig. 4), the reduction in axonal staining was more pronounced in LB males (Cohen’s d = 2.01) than in LB females (Cohen’s d = 1.27), and the intensity of staining was significantly correlated with contextual freezing (Fig. 6E). No significant effects of rearing, sex, or interaction were observed for the number of PDGFR1α progenitor cells or CC1 mature oligodendrocytes in the SLM (Additional file 1: Fig S5A-C). However, there was a significant reduction in MBP staining in the SLM of LB adolescent mice (F (1, 19) = 4.40, P = 0.049) that was again more pronounced in males compared to females (Additional file 1: Fig S5D). Together, these results indicate that reduced axonal staining and myelination persist in the SLM of adolescent LB mice and may contribute to the sex-specific deficits seen in contextual fear conditioning.
Fig. 6
LB causes more severe deficits in contextual fear conditioning in adolescent P34 male mice. A Experimental timeline. B Contextual fear conditioning. CTL male = 17, CTL females = 18, LB males = 13, LB females = 18. C, D NF-H axonal staining in the SLM of adolescent mice, N = 6 mice per rearing and sex group. E The intensity of axonal staining correlates with contextual freezing behavior. Error bars represent the mean ± SEM. *p < 0.05, **p < 0.01, ***p < 0.001
LB reduces reelin-positive projections in male miceMost axonal terminals in the SLM originate from the entorhinal cortex [27], suggesting that reduced axonal staining in the SLM (Fig. 6D) reflects abnormal connectivity between these two brain regions in LB adolescent mice. Indeed, dMRI tractography revealed a significant reduction in structural connectivity between the entorhinal cortex and the dorsal hippocampus on the left (F (1, 20) = 8.241, P = 0.0095, Fig. 7A, B) and the right hemispheres (F (1, 20) = 5.659, P = 0.0274, Fig. 7C). Although there were no significant effects of sex or interaction between rearing and sex, the effect sizes in males were larger than those in females (Cohen’s d left: M = 1.39, F = 0.92; Cohen’s d right: M = 1.65, F = 0.39).
Fig. 7
LB reduces structural connectivity between the entorhinal cortex (ENT) and the dorsal hippocampus (dHPC) in P29 mice. Representative images (A) and quantification of ENT-dHPC tractography in the left (B) and right (C) hemispheres. N = 6 mice per rearing and sex condition. Error bars represent the mean ± SEM. *p < 0.05, **p < 0.01
To further validate these findings, we administered a fluorescently labeled retrograde tracer, Alexa-555-CTB, into the left SLM of P28 CTL and LB mice, tested contextual fear conditioning at P34, and processed the mice a day later to assess the number of CTB-positive cells in the left entorhinal cortex (Fig. 8A). LB mice showed a significant reduction in contextual freezing (F (1, 23) = 6.80, P = 0.016) that was more pronounced in males than in females (Fig. 8B). No significant effects of rearing, sex or interaction were observed for CTB labeling at the injection site (Additional file 1: Fig S6). However, there was a significant interaction between rearing and sex for the number of CTB-positive cells in the LEC (F (1, 12) = 11.08, P = 0.0060). This interaction was due to a fourfold reduction in the number of CTB-positive cells in LB males (P < 0.0001, Cohen’s d = 3.69) that was not observed in LB females (P = 0.22, Cohen’s d = 1.69, Fig. 8C, D). Moreover, the number of CTB-positive cells in the LEC was highly correlated with freezing behavior (Fig. 8E). Together, these findings indicate that LB reduces connectivity between the entorhinal cortex and the dorsal hippocampus in a manner that correlates with deficits in contextual memory and is more pronounced in adolescent males.
Fig. 8
LB causes sex-specific deficits in lateral perforant pathway connectivity in P35 adolescent mice. A CTL and LB mice were injected with the retrograde tracer Alexa 555-CTB into the left SLM at P28, tested in contextual fear conditioning (P33-34), and perfused to assess the number of CTB-positive cells in the LEC at P35. B Contextual fear conditioning. C Schematic depiction of CTB injection at P28 and CTB labeling in the LEC at P35. D Number of CTB-positive cells in the LEC at P35. E The number of CTB-positive cells in the LEC correlates with contextual fear conditioning
Previous work has shown that reelin-positive cells project from the LEC to the dorsal hippocampus and that these projections are essential for contextual memory [30, 35, 36]. To determine whether the CTB-positive cells in the LEC were reelin-positive, we repeated the CTB retrograde labeling described in Fig. 8A using a second cohort of adolescent mice and replicated the sex-specific reduction in the number of CTB-positive cells in the LEC (Interaction: F (1, 13) = 10.76, P = 0.0060, Fig. 9A, B). We confirmed that all CTB-positive cells were also reelin-positive, regardless of rearing and sex (Fig. 9A, C), and found that the total number of reelin-positive cells in the LEC was reduced in LB mice, an effect that was more pronounced in males (rearing: F (1, 13) = 31.39, P < 0.0001, sex: (F (1, 13) = 25.50, P = 0.0002, interaction: (F (1, 13) = 3.399, P = 0.088, Fig. 9A, D). The percentage of reelin-positive and CTB-negative cells was significantly higher in LB males than in all other groups (interaction: F (1, 13) = 6.375, P = 0.025, Cohen’s d effect size males = 2.63, females = 1.19, Fig. 9A, E), indicating that most reelin-positive cells in the LEC fail to project to the dorsal hippocampus in LB males.
Fig. 9
LB reduces the number of reelin-positive projections in adolescent males. A Representative images of Reelin- and CTB-positive cells in the LEC (scale bars in A are 50 µm). Higher magnification of rectangular areas of merged images is shown on the right. B The number of CTB-positive cells is reduced in the LEC of male LB mice. C All CTB-positive cells are reelin-positive regardless of rearing and sex conditions. D LB reduces the number of reelin-positive cells in the LEC. E Effects of rearing and sex on the percentage of reelin-positive CTB-negative cells in the LEC
Comments (0)