Overview of MAMs proteomics
Proteomics is one of the important approaches to exploring membrane proteins comprehensively. By employing advanced 4D label-free mass spectrometry (MS) analysis, particularly enabling the identification of a large number of non-redundant proteins, we investigated changes in the proteome profile of MAMs isolated from HK-2 cells. MAM proteins differentially expressed were identified through bioinformatics analysis in normal and hypoxia/reoxygenation model of HK-2 cells. Our findings may underscore the vital role of MAMs in renal ischemic reperfusion injury (IRI), particularly in association with mitochondrial energy metabolism and key biological processes such as apoptosis, division, and the cell cycle. Notably, the crucial MAM protein MFN2 may play a pivotal protective role in renal IRI.
An initial proteomic study of MAM was published in 2011, identifying 991 unique proteins by using SILAC quantitation [39]. After that, a few of studies have been performed on MAM proteomics, such as the cerebral cortex of mouse in Alzheimer’s Disease model [40], retina and brain MAMs isolated from diabetes mellitus mice [41, 42],and so on. Currently, research on MAM proteomics has primarily concentrated on neurological disorders, with relatively limited exploration in other fields, particularly in the context of organ ischemic reperfusion injury. Our study focused on MAMs in the context of renal ischemic reperfusion injury, shedding light on alterations in protein composition and potential functional implications.
Some scholars have posited that the function of MAMs plays a crucial role in organ ischemic reperfusion injury, encompassing oxidative stress, endoplasmic reticulum (ER) stress, mitophagy, calcium overload, among other factors. Nevertheless, a notable research gap exists, as no studies have systematically analyzed the proteomic changes of MAMs in renal ischemic reperfusion injury. Utilizing mass spectrometry-based proteomic analyses, our study identified MAM proteins in human HK-2 cells, investigating changes following hypoxia/reoxygenation injury, a model simulating renal ischemic reperfusion stress. The data obtained in our study not only contribute to filling this research void but also lay the foundation for a hypothesis regarding the initial phase of kidney ischemic reperfusion injury.
Molecular function of MAM Proteins in normal and HR HK-2 cells
In the present study, we have identified 4489 non-redundant proteins in HK-2 cells MAMs with 3531 quantifiable proteins in both HR and normal MAMs, among which 688 proteins significantly altered in HR HK-2 ells with ultra-high quantitative confidence. Several proteins that have been well-studied to localize in the MAMs were detected, including ACSL4, ACAT1, ERP44, ERO1ɑ, erlin1, VAPB, CANX, MFN2 and GRP75 and VDACs. Nonetheless, PACS2, as a MAM marker, was not detected in our study, which is consistent with Zhang [39]. This observation could potentially be attributed to their comparatively limited prevalence within the MAM proteome, their suboptimal retention efficiency during MAM fractionation, or their potential interference with the detection of other proteins. Simultaneously, the expression of PACS2 within the renal tubules is relatively low [26].
Consistent with previously reported molecular functions of MAM fraction, our functional annotation of the MAM proteins suggest that MAMs is heavily involved in energetic metabolism process, such as ATP binding, GTP binding, GTPase activity, ATPase activity. In our study, both total protein and differentially expressed proteins consistently point to the close association of MAMs with mitochondrial energy metabolism. This manifest the strong association between mitochondrial energy metabolism and renal ischemic reperfusion injury, a connection that has been validated in numerous studies [43,44,45].Considering the kidney's role as one of the organs with the highest oxygen consumption and the fact that IRI results from damage caused by energy deprivation and subsequent reperfusion, our findings suggest that MAMs play a crucial role in both the baseline energy requirements of the kidney in physiological states and under the pathological conditions of IRI. Oxidoreductase (ERO1ɑ, ERP44 et al.) and chaperone (GRP75, CypD) are two kinds of proteins that have been substantiated by a few studies to play an important role in MAMs, ranking second and third respectively in their significance (p = 5.94E−41, 1.08E−40 respectively) in total protein, although not being the most abundant proteins enriched within these structures. In a similar vein, the functional enrichment analysis revealed that oxidoreductase activity and chaperone binding remain prominently enriched among differentially expressed proteins. Oxidoreductases, implicated in electron transfer, serve as potential indicators of alterations in cellular redox balance—a phenomenon intricately linked to both oxidative stress and mitochondrial function. Concurrently, the observed enrichment of chaperones, critical for protein folding and stability, signifies their integral role in preserving cellular homeostasis.
However, it's noteworthy that the molecular components within this molecular function differ between total proteins and those exhibiting differential expression. This suggests that these activities might operate at different levels or through distinct mechanisms in physiological and pathological conditions. These findings underscore the dynamic and context-dependent nature of cellular processes involving MAMs, shedding light on their multifaceted roles in kidney physiology and pathology.
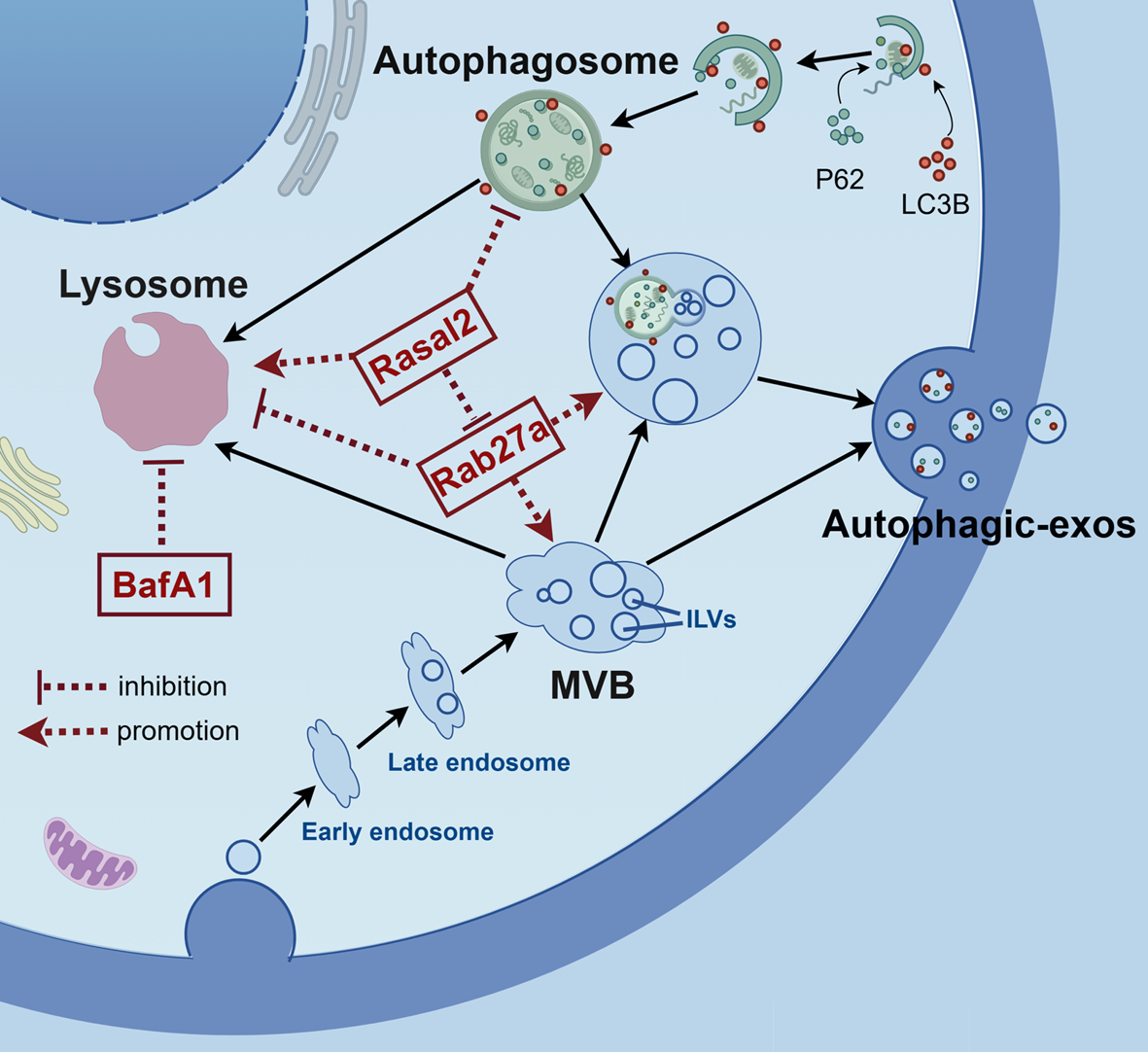
Statistically validated KEGG analysis unveiled that the most significant pathway was metabolic pathways in both total and differential proteins. Furthermore, several other vital metabolism-related pathways were also found to be involved, such as oxidative phosphorylation, carbon metabolism, the citrate cycle. The results suggest that mitochondria, as the most energy-productive organelles in eukaryotes, play an important metabolic role in MAM function. Not surprisingly, several KEGG pathways are related to the nervous system disease, such as pathways of neurodegeneration, amyotrophic lateral sclerosis, Alzheimer disease, Parkinson disease. The possible reasons are as follows: On the one hand, the MAM proteins are highly conserved between different species and different tissues; and on the other hand, the function of MAMs has received high attention in neurological diseases, which has been a lot of evidence to support this. As compared to brain research, ischemic reperfusion injury has fewer studies involving MAM proteins, leaving a huge research gap. It is essential to note that the mitophagy pathways, exclusively enriched in the analysis of differential proteins, encompass BCL2L13, BNIP3, ATG9A, MAP1LC3B2, EIF2AK3, RRAS2, MFN2, and RPS27A. These proteins exhibit potential involvement in mitophagy processes, a pivotal cellular mechanism responsible for selectively removing damaged or dysfunctional mitochondria. Each protein may contribute to mitophagy through distinct mechanisms: BCL2L13 engages in the crosstalk between apoptosis and mitophagy; BNIP3, a key regulator, facilitates recognition and clearance of damaged mitochondria; ATG9A contributes to autophagosome formation; EIF2AK3 links mitophagy to the unfolded protein response (UPR); MFN2, beyond its role in mitochondrial dynamics, coordinates mitophagy; and RPS27A, implicated in ubiquitination, may mark mitochondria for degradation through the ubiquitin–proteasome system. This exclusive enrichment underscores their potential significance in the cellular response to hypoxic stress, emphasizing their multifaceted roles in renal ischemic reperfusion injury.
Within the 688 differentially expressed proteins between the HR and N groups, the top five upregulated molecules were BNIP3, SEMA4B, RALGAPB, EPHB1, and SNAPIN, which may signify efforts to enhance cellular resilience and combat stress-induced damage. Conversely, the top five downregulated molecules were SAR1A, EIF3M, MFN2, PWP2 and PHF5A respectively, which may point to potential vulnerabilities in cellular processes. Among them, BNIP3 is associated with potential engagement in mitochondrial autophagy; SEMA4B may contribute to altered cell migration and apoptosis; RALGAPB implies regulatory involvement in cellular processes through GTP hydrolysis; EPHB1, as a tyrosine kinase, may signify engagement in signaling pathways crucial for cellular adaptation; SAR1A, EIF3M, and PWP2 may affect protein transport and synthesis, respectively; PHF5A may affect mRNA splicing; MFN2, being a multifunctional protein, has been examined in our subsequent investigations. These findings provide insights into the specific genes that exhibit significant changes in response to ischemic reperfusion injury.
As previously mentioned, the spliceosome has exhibited significant relevance both in biological processes and KEGG pathways. This suggests a potential close relationship between the spliceosome and MAMs, particularly in the context of ischemic reperfusion injury. However, there is currently limited research on the direct connection between the spliceosome and MAMs. Intriguingly, in a study of testis proteomics, the spliceosome also emerges as highly significant in the KEGG pathway, although the author does not delve extensively into this aspect. It's worth noting that a latest study has shed light on two splice variants of MFN2, namely ERMIN2 and ERMIT2, located on the endoplasmic reticulum (ER). These variants have been found to play crucial roles in maintaining communication between the ER and mitochondria, regulating endoplasmic reticulum morphology, facilitating calcium ion transport, and more [46]. This discovery could potentially mark the beginning of a new research avenue investigating the intricate relationship between splicing process and MAMs.
MAMs are intricately regulated by the actin cytoskeleton, which plays a pivotal role in driving fission and mitochondrial dynamics. The actin cytoskeleton emerges as a major driving force for the functional interplay between the mitochondria and ER, exerting its influence on diverse cellular aspects, including maintaining cellular morphology, regulating organelle distances, modulating signal transduction, and sustaining overall cellular function. Conversely, it is noteworthy that actin polymerization, a fundamental process in the actin cytoskeleton dynamics, is energy-demanding [47]. Significantly, our protein quantification analysis in the context of HR in HK-2 cells revealed a total of 25 actin or actin-related proteins. Among these, four genes—ACTR8, ARPC2, CAPZA2, and INF2—were found to be differentially expressed, with ACTR8 being upregulated and the remaining three proteins downregulated. The dynamic regulation of the actin cytoskeleton is a critical aspect of cellular responses to stress, including hypoxia/reoxygenation (H/R) injury. In our proteomic analysis of HK-2 cells subjected to H/R, we identified alterations in four key proteins associated with actin dynamics: ACTR8, ARPC2, CAPZA2, and INF2. ACTR8, an essential component of the Arp2/3 complex, showed increased expression, suggesting heightened actin branching. Concurrently, ARPC2, another Arp2/3 complex subunit, exhibited downregulation, potentially impacting actin nucleation [48]. CAPZA2, a subunit of the F-actin-capping protein, displayed decreased levels, indicating a potential reduction in the capping activity of F-actin [49]. INF2, a formin protein involved in actin polymerization, showed decreased expression, suggesting alterations in actin filament formation [50]. These dynamic changes in actin-related proteins likely contribute to the cellular responses associated with cytoskeletal remodeling, migration, and adaptation to the challenges imposed by hypoxic/reoxygenation stress.
Notably, MAMs play a crucial role in regulating calcium homeostasis, a factor that should not be overlooked. Consistent with previous studies, we also have identified those calcium channel proteins, such as IP3R, GRP75, VDAC1, SERCA, CANX, and so on. Under pathological conditions, including IRI, the normal maintenance of the number and function of MAMs may contribute to coordinating ATP production and timely clearance of damaged mitochondria through mitophagy, thus alleviating renal ischemic reperfusion injury. However, the abnormal and sustained increase of MAMs may also aggravate calcium overload and the transfer of oxygen free radicals, thus aggravating renal ischemic reperfusion injury. The changes of MAMs in different stages and degrees may play different roles. How to maintain a moderate number of MAMs and maintain their normal function is also a problem that needs to be further explored and solved. In this report, there wasn't significant difference between two groups. We speculate that there may be a time lag between the changes in mitochondrial dynamics and the occurrence of calcium imbalance in different stages of renal ischemic reperfusion. What we observed pertains to the initial phase of ischemic reperfusion, and the alterations in mitochondrial morphology and function may occur prior to the onset of calcium overload. The sequence of occurrence of various pathophysiological processes needs more in-depth research.
Validation of BNIP3 and MFN2 and further exploration of the role of MFN2 in IRI
As previously mentioned, validation experiments for BNIP3 and MFN2 were conducted, and the results from both in vivo and in vitro experiments were consistent with the findings from the proteomics analysis. BNIP3 showed a significant upregulation, whereas MFN2 exhibited a significant downregulation. Both of them are located in the mitochondrial outer membrane. BNIP3 is believed to play a crucial role in ischemic reperfusion injury, primarily through the induction of mitophagy mediated by multiple pathways. HIF-1α-BNIP3-mediated mitophagy has been confirmed plays a protective role in IR of kidney, heart and cerebrum [51,52,53]. BNIP3, with its role in mitophagy, presents an intriguing avenue for therapeutic strategies aimed at clearing damaged mitochondria and alleviating injury. Therefore, the elevation of BNIP3 following HR may be a stress-induced self-protective mechanism in the HK-2 cells. Based on the aforementioned study, BNIP3 may represent a compelling target for therapeutic strategies focused on the clearance of damaged mitochondria and alleviation of injury.
MFN2, apart from being among the top differential proteins, also emerged as a hub gene according to bioinformatics analysis, emphasizing its central role. MFN2 is a multifunctional protein with the following primary functions: (i) As a GTPase, it plays a central role in mitochondrial metabolism; (ii) As a mitochondrial fusion protein, it participates in mitochondrial fusion and contributes to the maintenance and functionality of the mitochondrial network, including its regulation of mitochondria-associated ER membranes [54,55,56]; (iii) It is involved in the clearance of damaged mitochondria through the process of mitophagy [57]; (iv) It plays a role in controlling the unfolded protein response during endoplasmic reticulum (ER) stress [58, 59]. These diverse functions underscore the protective role of MFN2 in the IR injury. In our observations, we noted a significant reduction in MFN2 levels during ischemic reperfusion. Additionally, by using of two different techniques, confocal microscopy and transmission electron microscopy, we observed an increase in mitochondrial fission, mitochondrial swelling, and a perturbation in MAMs during IR. These phenomena may or at least partly be attributed to the decrease of MFN2. Furthermore, when MFN2 was overexpressed, we observed a reversal of the aforementioned mitochondrial damage and restoration of MAMs. Therefore, our results provide further confirmation of the protective role of MFN2 in the context of ischemic reperfusion injury.
To further explore the role of MFN2 in renal tubules and its underlying mechanisms and associated pathways, we conducted a proteomic study on normal HK-2 cells and OE-MFN2 HK-2 cells. KEGG enrichment analysis revealed a significant positive correlation between the PI3K pathway and MFN2. Subsequently, we performed a correlation analysis between upregulated genes in the PI3K/AKT pathway and MFN2. The genes identified in the PI3K/AKT pathway included LAMA5, ITGB3, PRKCA, PPP2R5C, OSMR, THBS1, and TLR2. LAMA5, ITGB3, PRKCA, and PPP2R5C have been implicated in various cellular processes, such as cell adhesion, cell transformation, and cell cycle checkpoint regulation. However, their specific roles in the context of IRI remain limitedly studied. OSMR, regulated by the PI3K/AKT pathway, has been shown to exert protective effects in acute kidney injury (AKI) [60]. THBS1 is rapidly upregulated in renal IRI, exacerbating kidney damage by stimulating ROS production and inducing apoptosis in renal tubular epithelial cells. Inhibition of THBS1 has been demonstrated to alleviate renal injury [61]. TLR2, a member of the Toll-like receptors (TLRs) family, has been extensively studied in renal IRI and is recognized as a crucial initiator of inflammatory responses leading to renal injury and dysfunction in IR injury. TLR2 activates multiple pathways, including MYD88/NFκB, MAPK/JNK, PI3K/AKT, contributing to the production of proinflammatory cytokines, chemokines, increased ROS accumulation, cell apoptosis, and exacerbation of graft rejection [62,63,64]. In conclusion, the findings shed light on the potential significance of these genes in the context of renal IRI. Understanding their specific roles and interactions may pave the way for targeted therapeutic strategies aimed at mitigating the impact of IRI on renal function.
Subsequently, we validated the PI3K/AKT pathway. Western blot results were consistent with the proteomic findings, showing that MFN2 overexpression upregulated p-PI3K and p-AKT. Generally, activation of the PI3K/AKT pathway is considered protective IRI across different organs such as the kidney [65], brain [66], and heart [67]. The activation of PI3K/AKT provides protection through the upregulation of antioxidant, anti-inflammatory, and autophagy activities, inhibiting mitochondrial dysfunction and cardiomyocyte apoptosis. Numerous studies indicate that PI3K/AKT activation has a protective effect against myocardial ischemia–reperfusion injury (MIRI) [62]. Additionally, non-coding RNAs have been implicated in the regulation of oxidative stress by modulating signaling pathways [10, 68]. The PI3K/AKT pathway is involved in Nrf2 activation, and its inhibition weakens Nrf2 transcriptional activity. AKT activation enhances adaptation to oxidative stress by activating Nrf2-related antioxidant signaling [69, 70]. In summary, apart from its protective role through the regulation of mitochondrial dynamics, MFN2 may exert its protective effects in ischemic reperfusion injury by activating the PI3K/AKT pathway. In summary, MFN2 may emerge as a potential target for preserving mitochondrial and MAMs integrity during renal IRI. Translating these findings into clinical practice involves exploring pharmacological or genetic interventions to modulate MFN2 expression or activity. Additionally, interventions targeting MAMs in general, considering their central role in cellular processes, could offer broader therapeutic strategies.
Limitations
Firstly, in selecting the HK-2 cell line as a model for hypoxia/reoxygenation experiments, it is essential to acknowledge that ideal in vitro models that precisely mimic the complexity of proximal tubule cells are limited. It's important to emphasize that the use of HK-2 cells represents a technical compromise, and the findings should be interpreted with an awareness of the inherent limitations associated with this cellular model while HK-2 cells have been widely used in similar studies. We have incorporated additional in vivo experiments to strengthen the translational relevance of our findings. However, it is important to note that further in-depth in vivo proteomic studies are warranted for a more comprehensive understanding.
Secondly, based on the structural and component characteristics, MAMs are believed to be highly flexible and adaptable structures capable of recruiting various signaling molecules according to the different pathophysiological states. Ischemic reperfusion injury involves multiple pathological and physiological processes, and the primary mechanisms at may differ at different stages of injury. Our study focused on the initial phase of ischemic reperfusion injury, and as a result, it may not fully reflect the dynamic changes of MAMs at different injury stages. Adding more observation time points could potentially provide valuable insights into the relationship between MAMs and ischemic reperfusion injury more systematically and comprehensively.
Potentially further and futuristic research
The current study paves the way for future research in several promising directions. One potential avenue is to delve deeper into the functional roles of specific proteins or pathways identified within MAMs and their contribution to renal ischemic reperfusion injury (IRI). By elucidating the precise mechanisms by which these proteins or pathways operate in MAMs, researchers can gain a better understanding of their involvement in the pathophysiology of renal IRI, potentially leading to the development of novel therapeutic strategies. Additionally, exploring potential therapeutic targets within MAMs to mitigate renal IRI and associated diseases represents an exciting prospect. By identifying and targeting specific molecules or pathways within MAMs that are dysregulated during renal IRI, researchers may be able to develop targeted interventions aimed at preserving mitochondrial function and mitigating tissue damage in the kidney. Exploring the application of emerging technologies, such as CRISPR-Cas9 or single-cell proteomics, could provide valuable insights into the intricate mechanisms underlying MAM dynamics during renal IRI. Moreover, our study hints at a potential link between spliceosomes and MAMs, suggesting an intriguing connection that warrants further investigation. Exploring the interplay between spliceosomes and MAMs could uncover novel regulatory mechanisms underlying mitochondrial dynamics and function in the context of renal IRI.
Overall, these future research directions have the potential to deepen our understanding of the molecular mechanisms underlying renal IRI and to uncover new therapeutic targets for the treatment of kidney injury and associated diseases. The identification of key proteins, pathways, and interactions within MAMs presents opportunities for developing targeted diagnostic and therapeutic strategies. These could include biomarkers indicative of MAM dysfunction or interventions aimed at modulating MAM-associated processes to ameliorate kidney injury. We have also underscored the need for further translational research to bridge the gap between laboratory findings and clinical applications. Future studies exploring the diagnostic potential of MAM-related biomarkers or testing interventions targeting MAMs in relevant clinical settings would be instrumental in realizing the clinical implications of our work.






Comments (0)