
Remember me

Clinical descriptions of the cases and details of experimental methods are available in Supplementary materials and methods.
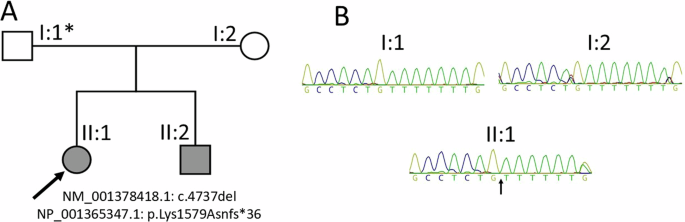
Nanopore T-LRS in both siblings was used to detect any additional variants (Fig. 1A). The target region, comprising 0.27% of the whole genome, was enriched with a bed file assigning DDC and its upstream and downstream 4-Mb buffer regions. The target region was successfully captured with a mean read depth of 39.02X (brother) and 29.29X (sister). Variant calling was performed using Longshot [8] and Clair with SNVoter [9]. After haplotype phasing, we searched for candidate variants in the other allele from the known variant (NM_000790.4:c.329 C > A, p.Ala110Glu) allele and identified a deep intronic variant (NM_000790.4:c.714+255 C > A) (Fig. 1B–D), which was absent in the gnomAD v3.1.2 and ToMMo 38KJPN databases. This genetic alteration is common in siblings and is predicted by SpliceAI [10] to activate the noncanonical splicing acceptor site (delta score of Acceptor Gain 0.25; pre-mRNA position −33 bp; delta score of Donor Gain 0.18; pre-mRNA position −213 bp). We then investigated by SpliceAid2 [11] the predicted effect of this variant regarding splicing regulatory factor motifs, revealing that UUUUAC, the binding site of Src substrate associated in mitosis of 68 kDa (SAM68), which regulates mRNA splicing [12, 13], was lost in the variant allele by becoming UUUUAA, causing abnormal splicing.
Fig. 1
Genetic analysis of DDC variant. A Family pedigree and DDC genotyping. The proband (II-1) and his younger sister (II-2) were born to non-consanguineous parents. Adaptive T-LRS was performed in the affected siblings. B Haplotype phasing analysis from Nanopore long-read sequencing data of II-1 visualized with the Integrative Genomics Viewer (IGV). DDC shown in reverse strand. Left panel: allele-phasing image showing sequence around chr.7:50537966 (hg38), c.329 C > A p.(Ala110Glu) variant, which is a known pathological variant present in haplotype-2 (HP-2). Right panel: sequence around chr.7:50527882 (hg38), c.714+255 C > A variant, which was in trans to c.329 C > A p.(Ala110Glu) and located in haplotype-1 (HP-1). C Sanger sequencing showing compound heterozygous variants in DDC (black arrow). The c.329 C > A variant was inherited from the father and c.714+255 C > A from the mother. D Genomic organization of DDC and location of variants identified in this study. Variants found in this study shown above and reported founder variant c.714+4 A > T shown below. UTR and coding region shown as light gray and dark gray rectangles, respectively
Based on the GTEx Portal, DDC expression was extremely low in lymphoblastoid cells (median TPM:0.01) and fibroblast cells (median TPM:0.00). Therefore, we performed a minigene assay to determine the impact of the c.714+255 C > A variant on transcript splicing (Fig. 2A). After transfection into HEK293, HeLa, and Neuro 2 A cells, the splicing patterns of the transcripts generated from the wild-type (WT) and variant (Mut) constructs were compared by RT-PCR and sequencing (Fig. 2B, C). Three WT transcripts were detected, but only one transcript of exons was produced from the Mut variant in all transfected cell types (Fig. 2B). We confirmed all the transcripts by Sanger sequencing: WT (upper: exon 6, intron 6 retention and cryptic exon 6a; middle: exon 6 and 6a; lower: only canonical exon 6) and Mut (only one transcript of exons 6 and 6a). The c.714+255 C > A variant activated a novel cryptic splice site at +288 to produce an aberrant splice isoform of DDC (Fig. 2C). This noncanonical 181 bp cryptic exon (exon 6a) introduced a premature stop codon, which could lead to nonsense-mediated mRNA decay (Fig. 2D, E). It was also observed in WT-transfected cells, but the same DDC mRNA isoform has previously been detected at low frequency (~1.5% to 21%) in normal cells [14]. Notably, the c.714+4 A > T variant with a founder effect in Taiwan and China, as the most common variant in AADC deficiency, showed approximately 16%–42% of the aberrant mRNA isoform, including exon 5-exon 6 + 37 bp insertion-exon 6a-exon 7-exon 8 [14]. These results suggest that the +288 cryptic splicing site functions at a leaky level in normal cells, but its insufficient expression leads to AADC deficiency; however, activation of this minor splicing site by c.714+255 C > A may result in a significant decrease in normal DDC expression. A heterozygous pathological variant c.329 C > A p.(Ala110Glu), which has previously been reported to reduced catalytic efficiency [15], was identified in our cases. It was assumed that compound heterozygous variants of c.329 C > A and c.714+255 C > A resulted in loss of DDC function. Both affected siblings showed severe phenotypes, but gene therapy using an AAV-hAADC-2 vector effectively improved their motor function, which was suspected to be affected by defective AADC function [16]. The current results were obtained by transient transfection assay using a minigene system, and further studies under conditions resembling in vivo systems are needed, such as knock-in of neuronal cells and/or model organisms or generation of induced pluripotent stem (iPS) cells derived from patients.
Fig. 2
In vitro minigene assay demonstrated an aberrant transcript specifically produced by the deep intronic variant of c.714+255C>A. A Schematic representation of H492 minigene vector. The vector contained two cassette DMD exons 18 and 20, and DDC exon 6 and the surrounding intron sequences (light blue) between these exons were cloned into the NheI and BamHI sites with a reference sequence (WT) or c.714+255 C > A (Mut). Red circle indicates variant site. WT, Mut, and empty vectors were transfected into HEK293, HeLa, and Neuro 2 A cells. RT-PCR was performed using the primers indicated by arrows. B Resolution of splicing RT-PCR products of minigene assay for DDC. Agarose gel electrophoresis showed 808 bp (upper), 521 bp (middle), and 340 bp (lower) products from the vector carrying the WT allele, but only a 521 bp product from the vectors carrying the Mut allele. C Sanger sequence electropherogram showing three types of transcripts produced by minigene assay. D Schematic illustration of alternative splicing patterns of pre-mRNA of DDC exon 6 with partial intron 6 using H492 minigene vector. Coding region indicated by rectangles. WT shown above and c.714+255 C > A (Mut) shown below the gene structure. In the WT, SAM68 binds to the UUUUAC motif; however, in the Mut case, the base substitution from C to A is expected to make it difficult for SAM68 to bind UUUUAA, leading to enhancement of the noncanonical splice site. E Schematic diagram of alternatively spliced mRNA including cryptic exon (yellow rectangle) and partial intron retention (light gray rectangle). Stop codon indicated by blue arrowheads. Nucleic acid sequences of canonical exon 6 indicated by capital letters
Recently, Himmelreich et al. reported 143 DDC variants found in 348 AADC deficiency patients, but even in this large study, only eight variants were involved in splicing abnormalities [17]. These eight pathogenic or likely pathogenic splicing variants include those within +6 from the donor site and −10 from the acceptor site [17]. It is assumed that unknown variants causing splicing abnormalities may exist in the deep intron of DDC.
Selective sequencing of targeted genomic regions has recently been developed and used to detect pathological variants in non-coding regions, and to confirm the diagnosis of autosomal recessive disorders in which variants in one allele cannot be found [18,19,20]. The deep intron variant found in this study could have been detected by short Whole-Genome Sequencing, but the sequence reads are too short to phase alleles harboring respective variants. Instead, T-LRS could detect the hidden variants very efficiently together with possible phasing of two variants.
The current study reports on siblings with a novel deep intronic variant of DDC leading to the inclusion of a cryptic exon associated with early truncation. It is promising that long-read sequencing with haplotype phasing can efficiently find hidden variants in unresolved autosomal recessive disorders.
Comments (0)