
Remember me
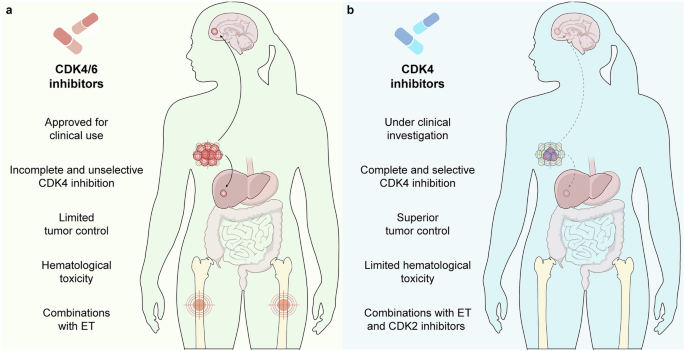



Two recent reports in Cell Research present cryo-electron microscopy structures of the Rpd3S HDAC complex bound to the nucleosome. These structures provide novel insight into the binding properties of the Rpd3S subunits Sin3, Rco1 and Eaf3 to DNA and Rco1 and Eaf3 to unmodified H3K4 and methylated H3K36, respectively, and also provide a fascinating look at the unique properties of Rpd3 in recognizing specific acetylated lysines for deacetylation in the context of Rpd3S.
The enzyme Rpd3 from the budding yeast Saccharomyces cerevisiae is a member of the Class I histone deacetylases (HDACs). This group of HDACs is highly conserved throughout eukaryotes. The most widely recognized role of Rpd3 and many other HDACs is as an antagonist to the histone acetylation which is widely associated with active gene transcription.
Rpd3 has a well-established role in transcription repression by recruitment to gene promoters through sequence-specific repressor proteins. While this mechanism still figures prominently in gene regulation, initial works from our group and others showed a completely different role for Rpd3.1,2,3 These works established that Rpd3 exists in two, large (Rpd3L) and small (Rpd3S), multisubunit complexes. While Rpd3L was determined to carry out the long-established repressive role at promoters, Rpd3S was found to function within the gene body to maintain a deacetylated state of histones in the wake of the elongating RNA polymerase II (RNAPII). The Eaf3 subunit of Rpd3S contains a methyl-lysine binding region known as chromodomain (CHD). Rpd3S is recruited by the CHD to sites in the transcribed gene body with a histone H3 lysine 36 (H3K36) trimethylation modification by the Set2 histone methyltransferase that travels with the elongating RNAPII.
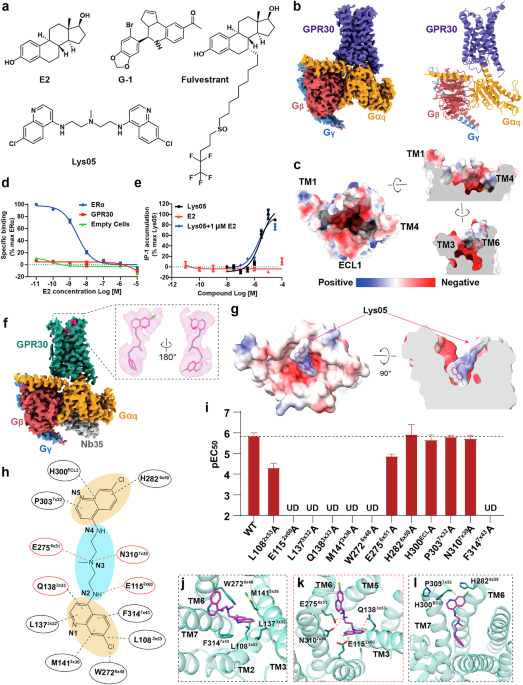
Two cryo-electron microscopy (cryo-EM) studies in Cell Research provide novel insight into the overall structure of the Rpd3S complex and how this complex binds its nucleosomal substrate to catalyze histone deacetylation.4,5 These two studies give views into the contacts between the Rpd3S complex, DNA and the H3 histone N-terminal tails, and the arrangement of these tails in the Rpd3 active site. The Rpd3 complexes have a core subunit composition found in both Rpd3L and Rpd3S that consists of the subunits Sin3, Ume1, and Rpd3. One of the first observations coming from both reports regards the subunit stoichiometry found in the Rpd3S complex. The core subunits Rpd3 and Sin3 are represented in a heterodimer configuration. Neither report included the Ume1 core subunit in their structure. Rco1 and Eaf3 are present in two heterodimers. This makes the Rpd3S a seven-subunit complex within the structures in these two reports (Fig. 1). This stoichiometry corresponds to previous non-structural biochemical works showing that Rco1 and Eaf3 were present as dimers in Rpd3S.6,7
Fig. 1: Rpd3S–nucleosome structures.
a, b Atomic models of the cryo-EM structure of the Rpd3S–nucleosome solved by Dong et al.4c, d Atomic models of the cryo-EM structure of the Rpd3S–nucleosome solved by Zhang et al.5 Important residues are highlighted in atomic sphere representations. Unmodified lysines, blue; methyl-lysine, green; lysine to glutamine substitution, orange; nonpolar/aromatic, yellow. Images were prepared using Chimera X molecular imaging software.11
Upon binding of Rpd3S to a nucleosome, both reports took a close examination of contacts between them. One of the features that establishes the uniqueness between these reports is the binding conditions used. Zhang et al.5 bound Rpd3S to nucleosome core particles (NCPs) that contained trimethylated histone H3K36 and 70 bp DNA exiting the nucleosome. By contrast, Dong et al.4 used a combination of different nucleosomes that contained or lacked a methyl-lysine analog of H3K36, an H3K9Q substitution that mimics an acetylated lysine, and/or 20 bp additional linker DNA entering and exiting the NCP. These different binding configurations by Dong et al. were designed to imitate different deacetylation scenarios.
Likely the two most important nucleosomal DNA-binding components in Rpd3S that emerged from these reports were found in Sin3, a core subunit, and Rco1, an Rpd3S-specific subunit. The Sin3 DNA recognition motif was mapped to an array of lysine residues in the Sin3 C-terminal domain that creates a basic surface to meet the negatively charged DNA (Fig. 1b, d). The DNA-binding activities of Rpd3S were tested by electrophoretic mobility shift assay. In addition to Sin3, lysine residues within the PHD domain of one of the Rco1 subunits bind the DNA exiting the nucleosome (Fig. 1a, c). This is supported by previous works showing that the Rco1 PHD is important for binding nucleosomes and suppressing intragenic transcription from cryptic promoters.7,8 Both reports also show that the Eaf3 CHD binds DNA adjacent to where the H3 N-terminal tails extend through the nucleosomal DNA (Fig. 1a–d). Dong et al. present evidence that Rpd3S exploits a linker tightening mechanism involving the lifting of the body of the Rpd3S complex while at the same time maintaining DNA contacts and rotating 150o relative to the nucleosome.
Beyond the shared observations regarding DNA binding of Sin3 and Rco1, both reports present different results that contribute to a broader picture of how Rpd3S interacts with the nucleosome. One prediction based on current functional models would be that Rpd3S binds trimethylated H3K36 in these structures. The structures assembled by Dong et al. were able to show that the CHD side chains are involved in binding the methyl-K36. They show that three aromatic residues within CHD form a shell surrounding the methy-K36 (Fig. 1b). Zhang et al. also show that the CHD localizes to H3K36me3 for one copy of Eaf3 (Fig. 1c). Interestingly, Zhang et al. demonstrate that the same Rco1 PHD domain that binds the exiting DNA also binds an unmodified form of histone H3K4 (Fig. 1c). In the gene bodies, H3K4 is unmodified, while at gene promoters H3K4 is trimethylated. This observation by Zhang et al. supports the current model in which Rpd3S through the Rco1 PHD domain recognizes the unmodified H3K4 in gene bodies, while Rpd3L through its Pho23 subunit PHD domain recognizes H3K4 trimethylation at gene promoters where transition of genes from an active to a repressed state occurs.7,9
The final highlight from these reports is observations regarding the recognition of acetylated lysine along the histone H3 N-terminal tail in the context of Rpd3S. In the structure described by Zhang et al., histone H3K18 was found to be positioned at the Rpd3 active site simply as a result of formation of the Rpd3S–nucleosome structure in their experiment. H3K18 along with H3K9, H3K14, H3K23, and H3K27 are known targets of acetylation on the H3 N-terminal tail. When Zhang et al. tested their Rpd3S–nucleosomes for deacetylase activity, they found that lower amounts of Rpd3S could achieve more efficient deacetylation on H3K23 and H3K14 as compared to H3K9, H3K18 and H3K27. Their conclusion was that the H3K18 acetylation is the most stable binding state, as it spontaneously occurred in their structure with low activity, and thus would have a low turnover rate. This is consistent with another study showing that Rpd3S had lower activity on H3K18 as compared to H3K14.10 By generating various nucleosomal binding partners for Rpd3S, Dong et al. were able to demonstrate various catalytic configurations for Rpd3S on the nucleosome. In addition, by using a nucleosome with a glutamine substituted at H3K9 which acts as an acetylated lysine mimic, they were able to form a structure with Rpd3S immobilized at H3K9 (Fig. 1a). This was the structure used for the majority of their binding observations. Based on the structures presented in these two reports, it is apparent that Rpd3S has the physical capacity to deacetylate the H3 N-terminal tail spanning K9 to K18. However, tests of its catalytic activity demonstrate varying efficiencies of deacetylation on these lysine residues. This raises an important question as to whether this reflects a biological role in specifying the lysines for Rpd3S deacetylation.
It is well-established that histone acetylation accumulates in the gene body in the absence of Rpd3S and this leads to cryptic transcription initiating from the gene body. In the presence of Rpd3S, these acetylation events must still occur but rapidly turn over. The exact lysine acetylations that are present when the nucleosome structure is restored in the wake of RNAPII are not clear. The variability in deacetylation efficiency by Rpd3S clearly suggests that the complex exhibits some specificity towards lysines in histone tails. Perhaps, this is a clue to the acetylation marks that are transiently placed on histones in the wake of RNAPII, and the mechanism that Rpd3 has evolved to rapidly turn over specific acetylated lysine residues.
Comments (0)