
Remember me
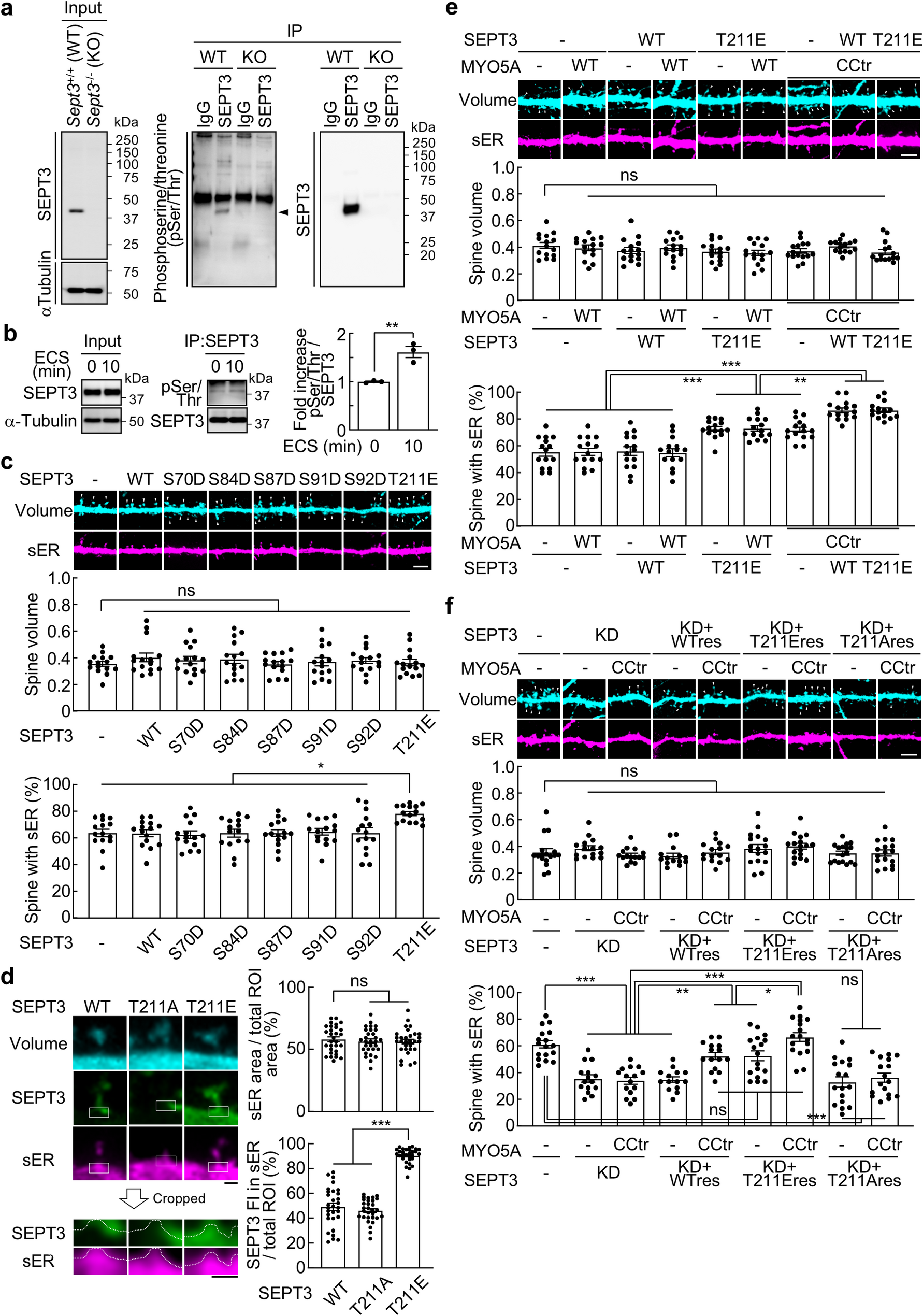






Synaptosome fractions were sonicated as described in methods. Sonicated and parallel unsonicated ‘control’ samples were layered on sucrose gradients (1 ml each of 0.32 M and 0.85 M sucrose) and centrifuged (50,000 rpm for 1 h in a Beckmann SW55 rotor) to remove soluble and light particulate material. The denser material pelleted through 0.85 M sucrose was collected and compacted through centrifugation in 20 mM HEPES and fixed for EM analysis.
EM examination showed PSD-containing structures in both control and sonicated pellets, either with or without presynaptic attachments (Fig. 1). In order to assess the relative number of clefts dissociated upon sonication, each PSD-containing structure was classified as either a PSD ‘with presynaptic elements’ (Fig. 1C) or a bare PSD ‘without pre-synaptic elements’ (Fig. 1D). Control pellets mostly exhibited intact synaptic junctions with PSDs apposed to presynaptic elements (89%, Fig. 1E). In sonicated pellets, on the other hand, only one in four of PSDs were with presynaptic elements (Fig. 1E).
Fig. 1
Effect of sonication on synaptic junctions. Synaptosome preparation (300 µg protein in 3 ml) is sonicated, pelleted through 0.85% sucrose, fixed and examined by electron microscopy. Unsonicated control samples (A & C) mostly exhibit intact synaptic junctions with apposed pre- and postsynaptic membranes (encircled), whereas in sonicated samples (B & D) PSDs without adjoining presynaptic elements (arrows in B) are predominant. PSDs in sonicated samples have associated postsynaptic membranes (D). Scale bars = 100 nm. Quantitative evaluation of data from one experiment, where parallel samples were scored for PSD-containing structures (control n = 80; sonicated n = 130) shows drastic increase in PSDs without presynaptic elements upon sonication (E)
Closer examination of the PSDs devoid of presynaptic elements in sonicated pellets revealed that they still retained the adjoining postsynaptic membrane (Fig. 1D). In an attempt to devise a strategy for the further purification of these structures, we reasoned that PSDs without the presynaptic elements (Fig. 1D) would have a lower membrane to protein ratio compared to intact junctions with both pre- and postsynaptic membranes (Fig. 1C) and therefore should be of higher density. Thus, a density-based fractionation protocol was implemented for the separation of PSDs from intact synaptic junctions.
SPM preparations (pellets from lysed synaptosomes) were sonicated and fractionated by density-gradient centrifugation in a discontinuous gradient consisting of 0.85 M and 1.2 M sucrose layers as described in Methods. Since synaptosomes and SPM fractions containing synaptic junctions are conventionally collected at the interphase above the 1.2 M sucrose, it was predicted that material with intact junctions would be retained at this interphase while the denser PSDs would pellet through. Lighter particulate material, on the other hand, is expected to be retained at the interphase above 0.85 M sucrose, while soluble proteins would remain in the supernatant in 20 mM HEPES buffer.
Pelleted material as well as material from the two interphases and the supernatant were collected (Fig. 2A) and analyzed for protein content. Figure 2B shows Coomassie stains of fractions recovered from the same amount of starting material. As expected, virtually all proteins from un-sonicated control samples were recovered at the interphase above 1.2 M sucrose layer, indicating preservation of the synaptic junctions. In contrast, proteins from sonicated samples were distributed throughout the gradient, reflecting breakdown of synaptic junctions (Fig. 2B).
Fig. 2
Purification of PSDs with associated postsynaptic membrane: fractionation of sonicated samples. Control and sonicated synaptic plasma membrane (SPM) preparations are fractionated by centrifugation through a sucrose gradient. Material is collected from four fractions: supernatant, interphases over 0.85 M and 1.2 M sucrose layers and pellet (A). Fractions corresponding to control and sonicated samples are analyzed by PAGE, Coomassie Blue staining (B) and Western Immunoblotting (C). Each lane in B and C corresponds to material recovered in indicated fraction from the same starting amount of SPM preparation
Immunoblots in Fig. 2C show the fractionation of specific synaptic proteins upon sonication. Again, lanes represent fractions recovered from the same amount of starting material. In accordance with the Coomassie protein profile, in control samples, all of the proteins tested were basically recovered at the 1.2 M interphase. On the other hand, in sonicated samples, the proteins became distributed among two or more fractions.
Upon sonication, major PSD scaffolds, PSD-95, SynGAP and Homer1, as well as components of AMPA and NMDA types of glutamate receptors, GluR1 and NR2A/B became most prominent in pellets (Fig. 2C, Supplementary Table 1). Considering that the pellet represents a much smaller proportion of the total protein recovered (Fig. 2B), the results indicate a high degree of enrichment of PSD-specific proteins [15] in that fraction.
On the other hand, presynaptic components [16], syntaxin, an integral membrane protein, and SNAP25, a membrane-associated presynaptic protein, fractionated predominantly into 0.85 and 1.2 M sucrose interphases, with relatively low levels in pellets. SNAP25 showed a wider distribution, suggesting some dissociation and redistribution during sonication. All of the proteins tested were also detected at the 1.2 M interphase, but at reduced levels, presumably corresponding to the minor pool of synaptic junctions that remain intact after sonication.
EM observation of glutaraldehyde-fixed pellets from sonicated samples (Fig. 3) showed PSDs with associated postsynaptic membranes from the top to the bottom of the cross-sectioned pellet. The high yield of PSDs without presynaptic elements upon sonication was verified in three preparations, with an average of 96 ± 1% of PSD-containing profiles without presynaptic elements [two experiments using standard SPM preparations: 95.5% (173/181), 93.4% (113/121), and one using an SPM preparation from older (24–48 weeks) rats: 96.6% (256/265)].
Fig. 3
Purification of PSDs with associated postsynaptic membrane. Examination of pellets from sonicated samples by electron microscopy reveals predominance of PSDs (arrows) with associated postsynaptic membranes but without presynaptic elements. Contaminants include membranous elements (asterisks) which are more prevalent in the top of the pellet, and mitochondria (m) which are more abundant in the bottom of the pellet. Scale bar = 1 μm
The most widely used protocol for the preparation of PSD-enriched fractions, devised by Cohen et al., [2] involves treatment of synaptosomal fractions with TritonX-100. Figure 4 allows a comparison of Triton-derived PSDs and sonication-derived PSDs from the present study. While PSDs appear to conserve their in situ morphology under either protocol, only PSDs separated through sonication retain the adjoining postsynaptic membrane (arrow in Fig. 4).
Fig. 4
PSDs separated by sonication (A) exhibit an associated postsynaptic membrane (arrow), whereas PSDs in the classical Triton-derived preparation (B) are devoid of postsynaptic membrane. Scale bar = 100 nm
Comments (0)