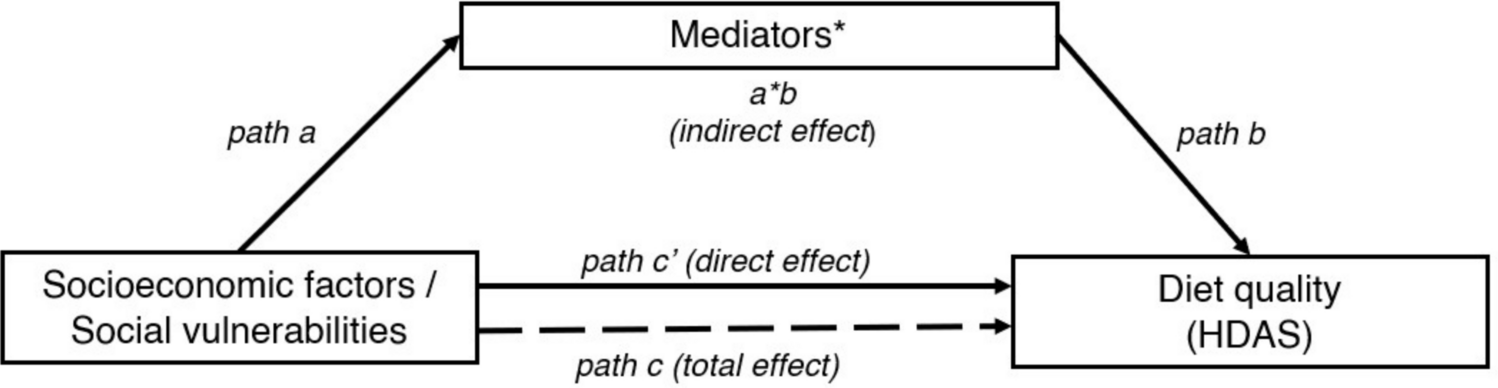
Existing research primarily focuses on the metabolic effects of carbohydrate restriction, either without energy restriction or with severe energy restriction in the form of fasting. Our study uniquely bridges this gap by directly comparing these dietary strategies, specifically isolating the effects of carbohydrate restriction on acute postprandial substrate and energy metabolism following food intake. We underscore the pivotal role of carbohydrate intake in modulating metabolic responses, independent of energy levels, and explore its potential to replicate the metabolic advantages associated with IER, without directly targeting weight loss. The findings provide new insights into dietary strategies showing that manipulating carbohydrate intake can achieve metabolic effects akin to acute fasting.
A key finding from our study is that LC diets elicited statistically similar metabolic responses across most measurements, including the primary outcome, postprandial TAG responses. Both LC diets achieved significant reductions in TAG levels, underscoring the role of carbohydrate restriction in attenuating postprandial lipaemia, a critical cardiovascular risk factor [4] independent of energy restriction.
Further analysis revealed similar responses in other substrate metabolites, with notable exceptions being fasting levels of 3-OHB and GLP-1, both primary indicators of ketogenesis and metabolic regulation, respectively. Notably, both LC25 and LCEB elevated 3-OHB levels, consistent with previous LCHF studies [32,33,34], though only LC25 reached significance. This may be due to LC25’s lower fat intake enhancing endogenous lipolysis and ketogenesis, while LCEB delivers more exogenous chylomicron-derived TAG [35, 36]. This may also be a consequence of, the overnight fasting being of insufficient duration to elevate ketone levels in LCEB, limited power due to sample size, and individual variability. Moreover, the failure to see statistically significant increases in fasting NEFA levels across both LC diets, as observed in other studies [18, 32, 33], may be due limited sample size. However, fasting levels of TAG, glucose and insulin levels remained unchanged, contradicting similar studies [18, 32, 33], potentially due to metabolically healthy participant profile. Nonetheless, upon consumption of a standard meal, chylomicron-TAG became the primary fat source [37] and hepatic 3-OHB production rose similarly in both LC diets, paralleling other lipaemia markers.
The postprandial response to the test meal further demonstrated a metabolic state similar to acute fasting, characterised by a shift favouring fatty acids and ketones over glucose as the primary energy source [38]. This shift is evidenced by decreased postprandial RQ levels, carbohydrate oxidation and increased fat oxidation, aligning with prior research [18]. Reduced glucose availability prompts the diversion of fatty acids from circulating lipoprotein-TAG, facilitated by increased lipoprotein lipase (LPL) activity in muscle, and hepatic storage towards beta-oxidation and ketogenesis [39]. Demonstrative of this mechanism, our postprandial response to test meal further demonstrated an increase in hepatic production of 3-OHB and NEFA levels, consistent with similarly designed studies [18, 34, 40]. Favourable reductions in postprandial TAG levels after LC diets, are conducive to reduced fatty acid availability for VLDL-TAG secretion and hepatic TAG re-esterification and increased chylomicron-TAG utilisation in the postprandial state due to increased fatty acid oxidation [37] as seen in comparable fasting study with LC25 arm [18]. This supports the notion that limiting carbohydrates to 50 g/day induces a lipolytic state that may continue into the next day, despite subsequent intake of carbohydrate and insulin release. This can be attributed to reduced carbohydrate intake, driven by elevated muscle LPL activity in response to limited carbohydrate availability, rather than reduced energy intake.
Improvements in the lipaemia profile coupled to higher postprandial glucose levels after both LC diets in our study, align with findings from previous LCHF [33, 34, 41] and fasting-mimicking studies [18, 40, 42]. However, unlike previous findings linking elevated glucose to decreased first-phase insulin release as a consequence of higher fasted NEFA levels [33, 34], we found no significant changes in fasted NEFA levels or in first-phase insulin secretion or other insulin sensitivity analysis. Our observed stability in postprandial insulin levels suggests that increased glucose may not always stem from altered insulin dynamics. The observed reduction in postprandial NEFA levels across LC diets likely reflects enhanced NEFA uptake in muscle, potentially due to the observed elevation in fat oxidation, or suppressed NEFA outflow from adipose tissue (i.e., lipolysis suppression), rather than insulin resistance. Further tracer studies would be required to confirm this mechanism. This is further complicated by the decreased GLP-1 levels observed after both LC diets, which is not seen in earlier research [33, 34, 40, 43]. Given GLP-1’s insulin stimulating function, alternative mechanisms could be influencing glucose regulation, warranting a need for further analysis, possibly through C-peptide measurements, to better decipher these results.
Nevertheless, studies identifying concurrent elevations in NEFA and glucose levels [18, 34, 42] align with our findings and offer insight into the mechanistic basis. This can be part of adaptive metabolism, wherein enhanced NEFA uptake and increased beta-oxidation reduce glucose uptake via the accumulation of metabolic intermediates (acetyl-CoA, NADH, citrate) [44,45,46]. Such intermediates interfere with carbohydrate utilisation by impeding GLUT4 translocation, inhibition of glycolysis, and lower pyruvate dehydrogenase (PDH) activity [44,45,46], illustrative of the intricate lipid-glucose metabolic interaction. Additionally, heightened intracellular NEFA levels can temporarily overload mitochondria, leading to dysfunction, excessive reactive oxygen species (ROS) production, and accumulation of lipotoxic intermediates, disrupting insulin signalling contributing to insulin resistance across tissues [47,48,49]. Numao et al. [34] attributed this elevation in NEFA and glucose levels to the high fat content of their LCHF diet; however, our research shows similar metabolic effects in LC diets both with high-fat (LCEB) and very low-fat (LC25) suggesting glucose scarcity as the primary driver. A notion supported by fasting studies [40, 42] and our group’s previous study [18], which demonstrates dose-response effects of both complete fasting and 75% energy restriction on postprandial metabolism, attributable to differences in dietary carbohydrate content. Current findings support the concept that acute metabolic challenges are indicative of short-term tachyphylaxis, (a rapid decrease in response to a metabolic stimulus), which may either attenuate or transform with prolonged dietary exposure. The observed impairment in glucose handling could, therefore, be potentially mitigated over time through metabolic adaptation which indeed is supported by studies on repeated cycles of carbohydrate restriction, both with energy restriction (e.g., IER) [14, 50] and possibly without energy restriction [51, 52]. However, both non-energy restriction studies, due to their exploratory nature, require cautious interpretation and further research. The latter study’s focus on women with breast cancer following a strict Mediterranean LC diet that may not mirror typical eating habits, affecting broader applicability and adherence in non-research settings. The former study, on the other hand, involved participants aged nine to thirty, and had varying diet restriction days between groups (intermittent LC with 7 days vs. IER with 4 days), potentially affecting outcomes. The longer carbohydrate restriction in the intermittent LC group might lead to more pronounced metabolic adaptations, such as increased fat oxidation or changes in ketone body levels and impact adherence and gut microbiota, complicating dietary approach comparisons.
The enhancement in fat oxidation from a one-day low-carbohydrate diet, irrespective of energy content, and the response to reintroduced carbohydrates can be seen as demonstrative of metabolic flexibility [53]. Augmented uptake of NEFAs by peripheral tissue, enhances fatty acid oxidation, minimises TAG accumulation, and improves insulin signalling in skeletal muscle, potentially offering significant health advantages [54]. A recent study by Antoni et al. [14] has shown that IER, inherently low in carbohydrate, leads to better postprandial lipid responses than CER with equal caloric intake and matched weight loss. This suggests that IER enhances metabolic flexibility by regularly activating a fat oxidation state. In contrast, benefits of traditional, CER’s benefits primarily stem from weight loss, which usually lacks a substantial shift to fat oxidation. Similarly, another study [55] highlights metabolic improvements in IER without weight loss, underlining the significance of alternating fuel oxidation in dietary approaches. Coupling these observations with our current study, it could be hypothesised that cycling between low-carbohydrate and regular carbohydrate intake might amplify this flexibility, thus improving metabolic health independently of weight loss [16].
Also, it is important to note that in terms of practical application, the primary motivation for adopting LC diets is often for weight loss [56], which by itself brings metabolic benefits [57]. A secondary objective of our study was to explore potential energy balance effects. The LCEB arm also was proposed to increase compliance by reducing hunger and increasing food availability, especially important if long-term repeated cycles of LCEB are adopted. Interestingly, in the appetite sensations while participants felt increased hunger on restriction days, consistent with prior fasting studies [40, 42], the hunger decreased after a test meal in both LC diet (i.e. with or without an acute energy deficit), potentially correlating with higher ketone levels as also evidenced by VLCD and ketogenic diets [58]. Moreover, variations in hunger did not translate into subsequent food intake as there was no difference in 48-hour ad libitum food intake for either diet. This indicated that the LC25 group maintained a 75% energy deficit with little compensation in intake, consistent with prior research [18]. Moreover, real-life adherence in the long-term might differ, low carbohydrate diets tend to reduce spontaneous food intake over time [59], suggesting that long-term energy intake under LCEB diets might still lead to an overall energy deficit, underscoring the need for further studies.
With regard to energy expenditure our study found no significant changes in short term REE or DIT consistent with findings from short-term energy-restricted diets [18, 40, 42]. While some literature indicates an acute increase in energy expenditure with low-carbohydrate and fasting diets due to heightened gluconeogenesis, protein turnover, TAG-fatty acid cycling, and ketogenesis [20, 60], long-term outcomes remain mixed. Doubly labelled water studies report variable effects on energy expenditure (EE) in prolonged low-carbohydrate diets, with some indicating increases [61, 62] and others showing no change [63]. This variation could stem from inaccuracies in RQ estimation, which might overestimate EE values when based solely on VCO2 [64]. In contrast, IER studies typically note adaptive decreases in REE [14, 65], suggesting different metabolic adaptations. It should be noted, however, that despite varying findings on energy expenditure, it’s crucial to recognize that these changes are likely too minor to significantly impact energy balance through adaptive changes in REE.
The main strength of the study was the novel comparisons between two low carbohydrate diets at different energy levels and two energy-balanced diets at different carbohydrate contents, from both a metabolic and energy balance perspective. The crossover design is a strength due to minimising individual variability. Yet we acknowledge that our study was limited to only examining postprandial responses following a single test meal, which may not fully capture the variability in postprandial metabolism related to meal composition and timing. We also acknowledge the limited sample size and metabolically healthy profile of our cohort which may explain why some postprandial results between LCEB and nEB showed only a trend, without statistical significance. A larger or metabolically compromised group might have yielded more pronounced differences.
Summary and future direction
In summary, our study demonstrated that carbohydrate restriction, both with and without energy restriction induces short-term changes in fasted and postprandial glucose and lipid metabolism, along with similar effects on appetite measurements and subsequent energy compensation in healthy overweight/obese adults. The lack of fasted and postprandial differences between our low-carbohydrate intervention arms raises critical questions but suggest a dominant role of carbohydrate restriction itself in inducing metabolic adaptations, potentially overshadowing the differences due to energy content. Therefore, the metabolic threshold for inducing these adaptations might be more achievable, than previously thought. Further research is warranted to investigate how metabolism and behaviours adapt to repetitive cycles of carbohydrate restriction at different energy levels and whether the observed results could replicate the impact of intermittent energy-restricted dietary regimens.



Comments (0)