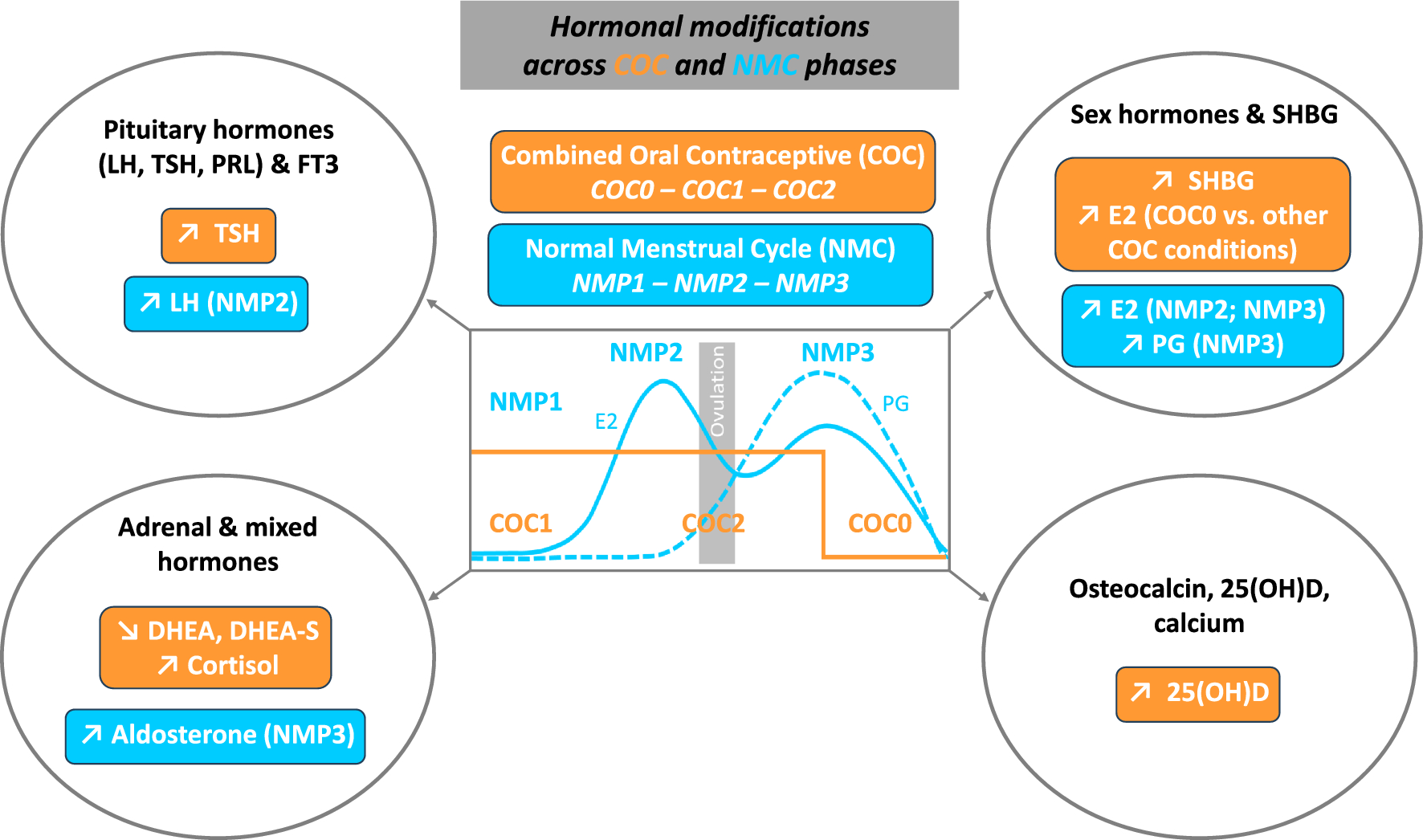
Given the scarcity of studies carried out on female athletes, we examined in this population the impact of COC (EE/LVN), across the different phases of active/inactive pill-taking, on various pituitary and peripheral hormones that play an important role in athlete performance and health. In parallel, we assessed in NMC athletes the effects of endogenous sex hormone fluctuations across the menstrual cycle on the parameters studied. Intake of COC vs. NMC, irrespective of the phase, increased TSH, SHBG, cortisol and 25(OH)D, and decreased DHEA and DHEA-S concentrations. Few parameters were influenced by the COC or NMC phases, with only E2 and aldosterone concentrations being higher during the washout and mid-luteal phases, respectively.
As anticipated (Castanier et al. 2021; Martin et al. 2018; Oxfeldt et al. 2020), in NMC subjects, we observed higher concentrations of LH during NMP2, with significantly elevated estradiol concentrations during NMP2 and NMP3 compared to NMP1. Progesterone, significantly higher during NMP3, was utilized to verify ovulation and thus the normality of the menstrual cycle in our NMC subjects. COCs logically inhibited ovulation, resulting in no LH peak nor PG increase, with significantly lower E2 concentrations in COC versus NMC subjects, except during the washout phase. Interestingly, there was a significant increase in E2 secretion during this hormone-free period, compared to the period of hormone intake, with similar values versus the early follicular phase. There is no consensus on this increase, or lack thereof, in E2 during COC0. This may be explained by the type of COC used and/or the timing of sampling. Indeed, our subjects were tested after an average washout of four days, following low doses of EE/LNG, unlike studies conducted on female athletes at the start of the inactive pill or with unspecified progestin nature (Ihalainen et al. 2021). In line with the literature (Coelingh Bennink et al. 2017; Knutsson et al. 2023; Özcan et al. 2023), we observed an increase in SHBG concentrations by more than 100% in our study with COCs, irrespective of the phase of COC administration. However, free estradiol index (E2/SHBG), considered to reflect estradiol activity (Gilberg et al. 2002), was not significantly different during the washout phase compared to the early follicular phase, in contrast to the first and second half of active hormone intake, where it was strongly decreased compared to NMC subjects. This suggests a relative increase in endogenous estrogenic activity during the COC-washout vs. COC-active hormone phases, which could partially compensate for the drop in exogenous hormone levels.
Prolactin and thyroid hormones each have pleotropic and independent roles allowing the body to accommodate to physical activity and exercise but they also share an interrelation in their responses, since hypothalamic thyrotropin-releasing hormone (TRH) release stimulates the release of TSH and thus the thyroids as well as the release of PRL, estrogen serving as an interconnective regulatory link by stimulating the release of both the thyroids and prolactin (Hackney and Saeidi 2019). Despite the variations of endogenous/exogenous estrogen concentrations across NMC and COC phases, no change in PRL levels was observed in the present study, either as a function of the menstrual cycle or hormonal contraception, and similarly, TSH values remained consistent irrespective of the menstrual cycle period. These results align with previous studies reporting little or no effect of the endogenous hormonal status in women (Aden et al. 1998; Kuhl et al. 1985a; Sawin et al. 1978; Weeke and Hansen 1975). However, we found increased TSH but not FT3 concentrations in our COC subjects, maintaining this increase during all COC periods. Higher TSH concentrations with COCs were initially described with concomitant higher total and free T3 and T4 concentrations (Kuhl et al. 1985b; Weeke and Hansen 1975), but most recent studies have reported an increase in TSH but not in free thyroid hormones, due to the parallel increase in thyroxine-binding globulin (TBG) protein (Özcan et al. 2023; Raps et al. 2014; Wiegratz et al. 2003). It may therefore be suggested that the discrepancy may be related to the lower doses of synthetic estrogens and progestins used in COCs today, limiting their thyroid impact.
With COC administration, we found a significant decrease in both DHEA and DHEA-S compared to NMC. This finding agreed with experiments performed on sedentary or recreationally active female subjects, with COCs including different progestins (Coelingh Bennink et al. 2017; Knutsson et al. 2023). COCs lower androgen status may drive mood and sexual function alterations in women although studies of these effects have yielded conflicting results (Burrows et al. 2012; Coelingh Bennink et al. 2017; Oinonen and Mazmanian 2002). Similarly, the higher total serum COR obtained here in the COC versus the NMC group was consistent with most previous works (Özcan et al. 2023). Unfortunately, we did not analyze cortisol-binding protein (CBP), but studies showed that COCs stimulate the liver production of binding proteins, including CBP, with, consequently, a probable lack of change in free cortisol concentrations and/or effects (Özcan et al. 2023; Vibarel-Rebot et al. 2015). Regarding NMC phases, according to the literature, we found no modification in the androgenic adrenal/gonadal DHEA and DHEA-S concentrations nor in COR throughout the menstrual cycle (Kanaley et al. 1992; Kuhl et al. 1985a; Knutsson et al. 2023; Vibarel-Rebot et al. 2015), whereas aldosterone levels were significantly increased in NMP3 compared with other conditions. There is very little data on aldosterone variations during the menstrual cycle, but one study (De Souza et al. 1989) previously noticed an increase during the mid-luteal phase. It may be hypothesized that this increase, whose involvement during endurance exercise needs to be determined, was mediated by the increase in its PG precursor, in view of the significant correlation obtained (r = 0.56).
Estrogens are considered as the “key regulator” of bone metabolism in women (Khosla and Monroe 2018) and changes in vitamin D and osteocalcin concentrations have been previously reported either across the menstrual cycle or with COC intake in healthy women or female athletes (Harmon et al. 2016, 2020; Jürimäe et al. 2011). Vitamin D is a fat-soluble steroid pro-hormone that occurs in two forms, vitamin D2 and D3, which may be obtained from sunlight exposure of the skin as well as from the diet. Both vitamin D2 and D3 undergo hydroxylation in the liver, where they are converted into 25(OH)D, further hydroxylated in the kidney to form 1,25-hydroxyvitamin D (calcitriol), a biologically active metabolite which then binds to vitamin D receptors (VDBP) at target tissues (Farrokhyar et al. 2015; Harju et al. 2022; Sist et al. 2023). Despite its well-recognized importance for both skeletal and non-skeletal systems, including cardiovascular, immune, inflammatory and muscle functions, a large proportion of the sporting population is deficient in vitamin D, with the risk increasing significantly in high latitudes, as well as in winter and spring, although specific prevalence in female athletes remains unknown (Farrokhyar et al. 2015; Harju et al. 2022; Sist et al. 2023). In our NMC group, 25(OH)D concentrations were below the recommended value of 30 ng/ml in most of our subjects (9/11), with two of them even showing vitamin D deficiency (< 20 ng/ml). These percentages (80%, 45% and 20% below 30, 25 and 20 ng/ml, respectively) appeared greater than those observed in sedentary or moderately active women (Harmon et al. 2020) and may reflect the increased needs resulting from intensive physical training. The same previous study only reported minimal change in calciotropic hormones during the menstrual cycle (Harmon et al. 2020) but, in women with 25(OH)D values below 30 ng/ml, lower mean E2 across the menstrual cycle. Partially in line with this study, we found similar 25(OH)D values across the menstrual cycle, probably because of its long half-life, but without any correlation with E2 levels. Interestingly, we observed in parallel relatively low calcium concentrations in our population and significant correlation between 25(OH)D and FT3 concentrations. There was surprisingly little information in female athletes (Nikolaidis et al. 2003) but it may be suggested that low blood calcium and FT3 may be partially resulted from low 25(OH)D values. Lastly, we found similar osteocalcin levels across the menstrual cycle or between our groups of subjects. This finding was partially in accordance with previous studies (Jürimäe et al. 2011; Martin et al. 2021) reporting no change across NMC but lower osteocalcin with COC administration (i.e., 20 mg of EE and 75 mg of gestodene) compared to NMC (Jürimäe et al. 2011). One hypothesis to explain the discrepancy with our COC data is the use of different progestins, whose specific impact on osteocalcin remains to be confirmed. However, our COC subjects had significantly higher 25(OH)D versus NMC subjects, with only one subject with vitamin D deficiency. This increase in 25(OH)D by COC intake was previously reported (Harmon et al. 2016) but as COC induced in parallel a 30% VDPB increase (Özcan et al. 2023), vitamin D status may be overestimated in COC subjects and the biological pathways involved remain to be determined (Harmon et al. 2016).

Comments (0)