
Remember me
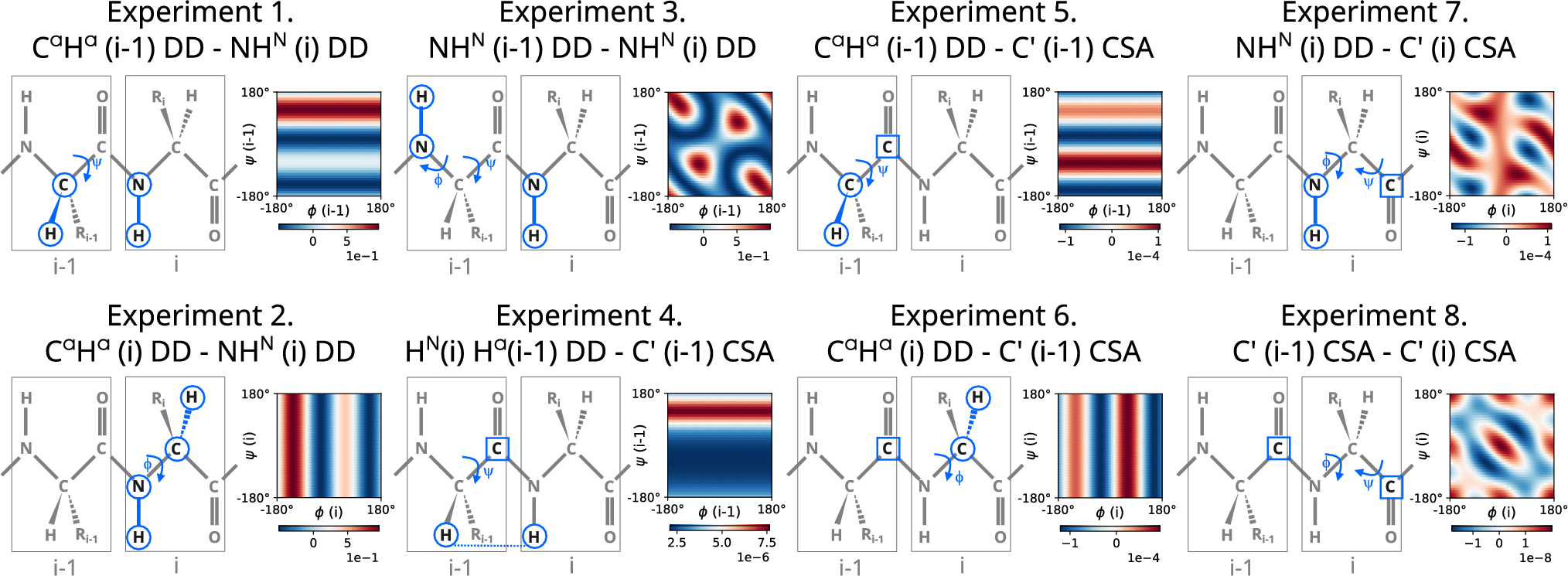
Relaxation times are determined by sampling a relaxing magnetization M after set of evolution times, τ (Kowalewski and Mäler 2006). For the pulse sequences commonly used to study proteins, an exponential decrease in signal intensity is observed. No special attention is paid to possible deviations from the exponential decay. However, a number of different effects interfere with measurements of the longitudinal relaxation rate R1 of amide backbone 15N nuclei in proteins: spin saturation of water proton affecting HN magnetization through exchange or NOE mechanisms (Grzesiek and Bax 1993; Chen and Tjandra 2011), incompletely suppressed DD/CSA interference (Boyd et al. 1990; Kay et al. 1992), or imperfect pulses (Lakomek et al. 2012; Ishima 2014; Gairì et al. 2015). Another source of perturbation in FID detection may be the non-ideal behavior of NMR transmitter and probe electronics exhibiting frequency-dependent interference (Uribe and Martin 2024). The mentioned above fast fading artifacts are difficult or impossible to eliminate or prevent. Consequently, the longitudinal 15N magnetization decay is no longer mono-exponential, and the initial experimental data points obtained for the shortest evolution times, τ, are the most disturbed. Consequently, the apparent R1 values are artificially increased and their standard deviations are enlarged. The anomalous effects become stronger at high magnetic fields. This phenomenon has usually been overlooked for two reasons. Deviations from mono-exponential decays can be hidden when spectra show a poor signal-to-noise ratio and/or R1 are derived from automatic procedures without careful inspection of the input data and intermediate results. Most of the artifacts can be quantitatively eliminated using a bi-exponential model rather than a mono-exponential decay model. However, the bi-exponential fitting procedure is prone to numerical instability due to strong correlations between the fitted parameters. Ishima (2014) recommended removing such artifacts by discarding the initial data points in mono-exponential fitting. She recommends using a τ values longer than 0.2 s for experiments performed at 21.1 T. However, the cutoff τ value can be expected to depend not only on the magnetic field strength but also on the setup of the individual NMR spectrometer and the properties of the specific sample.
To define more stringent conditions for R1 data processing, we measured R1 at 16.4, 18.8, and 22.3 T using a list of ten evolution times, τ (cf. Table S3). In addition to bi-exponential fitting, the R1 data were analyzed with mono-exponential decays, sequentially discarding the initial values of τ.
A visualization of the data obtained at 22.3 T for the 100% H2O sample is shown in Fig. 1. The R1 values determined for evolution times τ ≥ 0.32 s with a mono-exponential fit are equivalent to a bi-exponential fit. It leads to the conclusion that, in this particular case, the data for τ ≥ 0.32 s represent mono-exponential decay. On the other hand, too many rejected initial data points reduce the accuracy of the fit, as can be seen by comparing the data for 0.32, 0.48, and 0.64 s. Their average relative errors successively increase: 0.11%, 0.17%, and 0.26%. The site-specific differences between R1 values obtained from the bi-exponential and mono-exponential fits, calculated for different values of the initial τ value, display uniform distribution. Their mean value obtained for the shortest τ = 0.32 s is close to zero (Fig. 2). The numerical R1 values and their standard deviations shown in Fig. 2 are given in Table S1.
Fig. 1
Mean R1 values fitted to mono-exponential decay for increasing values of the shortest evolution time τ at 22.3 T (950-IH-AM: see the code description in the section NMR measurements). Error bars represent the average standard deviations of individual R1 values. The horizontal solid line represents the mean R1 value obtained by fitting data to bi-exponential decays and dashed lines represent the average standard deviations of individual R1 values
Fig. 2
Site-specific differences between mono-exponential and bi-exponential R1 values are calculated for initial τ value: 0.08 s and 0.32 s in the upper and lower parts of the Figure (code 950-IH-AM). Dashed lines correspond to the means of differences. Their numerical values for the upper and lower parts of the Figure are equal to 0.0144 s− 1 and 0.0003 s− 1, respectively
Similar results were obtained for the remaining measurements at 22.3 T (Figures S1 - S7). The bi-exponentially derived R1 values are on average 1% smaller than the R1s calculated mono-exponentially at 22.3 T for the 100% H2O sample. Consequently, the local mobility of the protein backbone can be erroneously estimated if evaluated from a model-free approach (Lipari and Szabo 1982) with inaccurate R1 values as input data (Chen and Tjandra 2011; Lakomek et al. 2012).
In contrast to the data obtained at 22.3 T, measurements obtained with the same method (HSQC, IBURP-2) for the 100% H2O sample at weaker magnetic fields (16.4 and 18.8 T) do not show a regular dependence of R1 value on the initial value of the evolution time τ (Figures S8 and S9) suggesting frequency dependent hardware problem. Such data can be safely processed using a mono-exponential decay model without discarding the shortest τ values. Three cases of the largest values of the Fisher-Snedecor statistics, F, in measurements taken at 16.4 and 18.8 T give the same R1 values for mono- and bi-exponential fits within the experimental error (Table 1). Therefore, the mono-exponential analysis of R1 is suitable for spectrometers with a magnetic field 18.8 T or weaker, at least for samples dissolved in H2O and the HSQC/IBURP-2 method.
Table 1 Two residues in the 800-IH-AM measurement and one residue in 700-IH-AM measurement show statistics F values greater than 5.143, corresponding to a probability level 0.05. The R1 values obtained from mono- and bi-exponential fits are within the experimental error limitsBased on our research, we recommend an initial check of the mono-exponential fitting model with subsequent rejection of the shortest evolution time data points. If the rejection of one or two initial data points does not regularly change the results, the full data set and the mono-exponential decay model can be used with caution. Otherwise, mainly for measurements in high magnetic fields, two options are possible: to use a bi-exponential fit of the R1 data, if the calculations are stable and the derived parameter errors are acceptable, or to discard the initial data points and then analyze a mono-exponential decay until no further changes in R1 values are observed.
Reproducibility of duplicated experimentsKnowing the reproducibility of the individual experiments is crucial for investigating very small effects, such as, for example, possible differences between HSQC and TROSY detection or differences between interference suppression methods. For this reason, several types of experiments were duplicated at intervals of several months. The statistical analysis of these measurements is given in Table 2.
Table 2 List of duplicate experiments with parameters characterizing pairwise differencesThe mean differences between duplicate experiments, <diff>, do not exceed 0.01 s− 1, and in many cases are even an order of magnitude smaller. The rms values of the differences are also less than 0.01 s− 1 with the exception of one case of 950-TH-HP experiments. These values limit the meaningless differences between the two experiments.
However, it should be born in mind that the changing the temperature of a sample usually results in a significant, often unrecognized source of experimental error. To assess temperature-induced errors, the temperature gradient of R1 was determined. Its value was equal to 0.028±0.003 s− 1/K at 16.4 T (details of the determination of the temperature gradient are given in the Supplementary Material). Given the maximum detected temperature deviation of 0.3 K in our work, temperature can cause R1 to vary by up to 0.008 s− 1. All < diff > values given in Table 2 do not exceed this value.
Comparison of HSQC and TROSY techniques applied to R1 measurementsTwo direct comparisons of HSQC and TROSY techniques can be found in the literature. Zhu at all. (2000) compared relaxation measurements of holo-calmoduline (17 kDa) sample at 17.6 T (750 MHz) removing interference effects during relaxation evolution times using 1H 180° HN-band selective pulses at 5–10 ms interval. The authors found that excellent agreement was obtained between TROSY-based experiments and the corresponding HSQC-based experiments. Lakomek et al. (2012) measured the small perdeuterated GB3 protein (6.2 kDa) at 14.1 T (600 MHz) using HN-band selective IBURP pulses every 40 ms to suppress interference effects during relaxation evolution times. These authors also found that no systematic differences were observed between R1 relaxation rates measured for GB3 using TROSY-detection and those obtained using HSQC-detection.
We conducted systematic comparison experiments based on TROSY and HSQC on two samples (100% H2O and 90% H2O/10% D2O) using three different interference suppression schemes at high magnetic fields of 22.3 T (950 MHz) and 18.8 T (800 MHz).
Table 3 Statistical analysis of differences between pairs of R1 relaxation values obtained by pairs of methods based on TROSY and HSQCAll twelve pairs of experiments given in Table 3 show very good statistical agreement between the TROSY and HSQC-based methods. The result shows that the two techniques produce virtually identical results, and the differences between them are comparable to those in the duplicated experiments.
Site specific differences for the 800/TH-AM/IH-AM pair are shown in Fig. 3. Another example is shown in Fig. S10. Well-defined ranges of anomalies cannot be recognized.
Fig. 3
Site-specific differences between TROSY-based and HSQC-based experiments at 18.8 T (800 MHz); codes 800-TH-AM and 800-IH-AM. The band HN selective pulses were used for interference suppression; solvent − 100% H2O. Solid line represents mean value of differences and dashed lines correspond to the mean ± 3σ. All differences are within this range
DD/CSA interference removal in R1 measurementsInterference effects between dipolar and chemical-shift anisotropy relaxation interactions in the measurement of relaxation times can cause significant errors in a simple mono-exponential analysis of the raw data (Goldman 1984; Werbelow 1996). Initially, interference was neglected in the analysis of 15N protein relaxation data (Kay et al. 1989; Clore et al. 1990). Shortly thereafter, Boyd et al. (1990) showed that broadband decoupling of 1H nuclei applied during the evolution delay, τ, removed interference effects. Kay et al. (1989) introduced a different approach, using 1H 180° pulses every 5–10 ms. For the staphylococcal nuclease protein (18 kDa), they observed an increase in R1 relaxation rate up to 9%. Moreover, the errors increase rapidly with decreasing molecular weight of the molecules studied. 1H 180° pulses were also used for the suppression of interference effects in the determination of transverse relaxation rates (Palmer III et al. 1992).
Another source of error in R1 measurements is due to the varying number of non-selective 180° 1H pulses used to eliminate relaxation interference effects during evolution times. Progressive suppression of water magnetization due to an increasing number of 180° 1H pulses for longer relaxation delays, τ, manifests itself in an increase in apparent R1. Elimination of this experimental defect has been achieved by replacing non-selective 180° 1H pulses with flip-back pulses (Grzesiek and Bax 1993; Chen and Tjandra 2011,) or HN-band selective pulses (Geen and Freeman 1991; Chill et all. 2006; Lakomek et al. 2012).
Gairí et al. (2015), studying small GB3 protein in two magnetic fields (14.1 and 18.8 T), observed that HN-band selective pulses (IBURP-2) resulted in smaller R1 values than the cosine-modulated IBURP-2 method, which selectively inverts two spectral bands, amide and aliphatic protons. The authors concluded that the differences in R1 values obtained with the different pulse trains were small but systematic and significant (rms diff = 0.075 s− 1) and greater than the reproducibility of the individual experiments (rms diff = 0.030 s− 1). They concluded that the source of this discrepancy could be either ineffective interference suppression or scheme-dependent saturation effects differently affecting HN magnetization at the start of each scan. Differential saturation effects should be sensitive to the recycle delay, whereas not fully effective interference suppression should not be affected by the delay. Rates of 15N R1 were measured using IBURP-2 and cosine-modulated IBURP-2 at recycle delay values of 1.7 and 3.5 s. The mean relaxation rates R1 measured by the cosine-modulated IBURP-2 method did not depend on the length of the recycle delay and were very similar to those obtained using IBURP-2 pulses with a longer recycle delay, whereas this method with a shorter recycle delay resulted in smaller values of R1. Therefore, the authors concluded that the cosine-modulated IBURP-2 pulses were superior to the IBURP-2 approach. However, in the experimental part the authors stated that cosine-modulated IBURP-2 pulses were 1.26 times longer and required four times more power (6 dB) than IBURP-2 pulses. It can be expected that stronger microwave heating without temperature compensation can increase the relaxation rate R1 based on the cosine-modulation of IBURP-2 compared to R1 derived from IBURP-2.
Our results include measurements of two ubiquitin samples (100% H2O and 90% H2O/10% D2O) at three magnetic field strengths, 16.4, 18.8, and 22.3 T. The interference suppression by the HN-band selective pulses (AM) almost never shows lower mean relaxation rates < R1 > compared to < R1 > measured with cosine-modulated IBURP-2 pulses (AMAL) (Fig. 4 and Figs. S11 and S12). The mean rms difference calculated for the 10 pairs AM/AMAL measurements is 0.012 s-1 (Table 4), while the reproducibility of the 10 experiments gives a comparable mean rms difference of 0.009 s-1. Differences between the two suppression methods are comparable to the reproducibility of the experiment and several times smaller than those observed by Gairí et al. (2015). The largest differences compared to Gairí’s data are: rms difference (Gairí) = 0.075 s-1, rms difference (22.3 T) = 0.010 s-1, rms difference (18.8 T) = 0.015 s-1, and rms difference (16.4 T) = 0.014 s-1. These can be attributed to temperature fluctuations and stronger microwave heating in the cosine-modulated IBURP-2 method.
Fig. 4
Mean R1 values measured with HSQC or TROSY based sequences at B0 = 22.3 T. DD/CSA interference was suppressed by IBURP-2 (AM), cosine modulated IBURP-2 (AMAL), or nonselective 180° (HP) pulses. Error bars represent average standard deviations of individual R1 values. Boxes mark corresponding AM/AMAL pairs. Dashed lines represent averages of all data points in appropriate parts of the figure. Difference of R1 values between AM and AMAL approaches observed for the mixed H2O/D2O solvent, <ΔR1 > = 0.02 s− 1, rms diff = 0.01 s− 1, might be attributed to 0.5 K temperature divergence
In addition, we determined R1 relaxation data using the IBURP-2 method for 100% H2O sample at 22.3 T with 3 recycle delays: 2, 4, and 6 s. The averaged R1 values are virtually identical (Fig. 5). Therefore, the difference between IBURP-2 and the cosine-modulated IBURP-2 cannot be explained by differential saturation effects. In conclusion, the IBURP-2 method seems to be superior to its cosine-modulated counterpart.
Fig. 5
Averaged R1 values determined with IBURP-2 (AM) approach for 100% H2O sample at 22.3 T with 3 recycle delays: 2, 4, and 6 s. Rms diff values calculated for pairs 2s/4s and 2s/6s are both equal to 0.005 s− 1. These rms diff values correspond to the values obtained for duplicated measurements
Table 4 Pairwise statistical analysis of differences between relaxation values obtained for interference suppression by the HN-band selective pulses (AM) and cosine-modulated IBURP-2 pulses (AMAL)One more observation reported by Gairí et al. (2015) requires comment. They found that the use of non-selective 180° 1H pulses, causing strong water saturation, resulted in an overestimation of R1. Furthermore, errors from water saturation effects varied along the protein sequence, being larger in the most solvent-exposed regions of the GB3 protein.
A close inspection of Fig. 4, S11, and S12 reveals, that suppressing interference with non-selective 180° 1H pulses has no effect on the averaged R1. Incidentally, this is a good example of how the the analysis of R1 averaged differences often gives a deceptively simple but false picture. Analysis of individual, residue-specific differences reveals a non uniform pattern. Site-specific differences between measurements using cosine-modulated IBURP-2 or non-selective 180° pulses for 100% H2O sample in three magnetic fields were calculated twice. In Fig. 6 all R1 values were obtained in a way that rejects fast fading artifacts, i.e. the data with the shortest evolution times τ = 0.32 s were used in the fitting procedures. The differences show no significant variation along the protein sequence for the use of non-selective 180° 1H pulses. On the other hand, in Fig. 7 all R1 values were obtained without discarding these artifacts. The highlighted rapidly exchanging HN residues are different from the other residues.
Fig. 6
Site specific differences between measurements applying the cosine-modulated IBURP-2 and nonselective 180° pulses for 100% H2O sample at three magnetic fields. The shortest evolution time τ = 0.48 s. Negligible rms diff values are equal to 0.006, 0.012, and 0.010 s− 1 for 22.3, 18.8, and 16.4 T, respectively. Solid lines represent mean values of differences and dashed lines represent the average standard deviations of individual R1 values
Fig. 7
Site specific differences between measurements applying the cosine-modulated IBURP-2 and nonselective 180° pulses for 100% H2O sample at three magnetic fields. The shortest evolution time τ = 0.08 s. Rms diff values are equal to 0.008, 0.012, and 0.010 s− 1 for 22.3, 18.8, and 16.4 T, respectively and do not differ from those given in Fig. 6. Amino acid residues with fast HN exchange are shown in red squares. Solid lines represent mean values of differences and dashed lines represent the average standard deviations of individual R1 values
Such an effect can be observed in certain situations, and it is discussed in the following section. It should be noted, however, that the data shown in Figs. 6 and 7 were obtained for a sample of 100% H2O. The differences between the samples of H2O and H2O/D2O are analyzed in detail in the following section.
Exchangeable deuterons perturb the apparent longitudinal relaxation rates 15N for solvent-exposed amidesMagnetic field stability is one of the key factors in NMR spectroscopy. For this purpose, a deuterium lock system is generally used. To make it work, a mixed H2O/D2O solvent (usually 90%/10% or 95%/5%) is used instead of pure H2O for NMR measurements of proteins. As the protons of HN exchange with both protons and deuterons of the solvent, a reduction in the signal intensity of HN is observed due to partial H→D substitution. This effect reduces the sensitivity but does not affect the R1 values. On the contrary, other mechanisms mentioned in the section “Elimination of systematic errors…” modulate the longitudinal relaxation. In magnetic relaxation measurements, HN exchange leads to erroneous values of the apparent relaxation parameters, provided that these exchange rates are not significantly slower than the relaxation rates.
There are nine amino acid residues in ubiquitin molecule that show increased rates of HN exchange: L8-T12, A46, and L73-G75 (Cordier and Grzesiek 2002). Moreover, these residues show reduced temperature gradients of the longitudinal relaxation rate R1 (Fig. 8). Such residues can be identified by a strong deviation from the mono-exponential fit of the experimental decay of R1. There are two methods to identify such residues.
Fig. 8
Experimental exchange HN values, k(HN) taken from Cordier and Grzesiek (2002); lower part. Temperature gradients of R1 relaxation rates (upper part) determined from the ubiquitin spectra measured at 288 and 308 K and 16.4 T; solvent 90% H2O/10% D2O, method HSQC, interference DD/CSA suppressed with nonselective 180° 1H pulses
Firstly, the mono-exponential determination of R1 values with and without the shortest evolution times, τ, which are most strongly disrupted, was analyzed. This perturbation of R1 values clearly appears in samples containing some D2O. It disappears in samples without D2O, as can be seen in Fig. 9 and S13. However, it can be, partially preserved in 100% H2O when non-selective 180° 1H pulses are used to suppress interference (Figures S14 and S6).
Fig. 9
Both plots display residue specific differences between R1 values determined with data comprising evolution times τ ≥ 0.08 s and data after rejecting the four shortest τ values (τ ≥ 0.48 s). Upper part - solvent 90% H2O/10% D2O, lower part - solvent 100% H2O. Interference suppression - HN selective band (AM). Similar comparisons for other field strengths and other techniques are presented in Supplementary Materials
Secondly, the bi-exponential fits of the experimental data, which show the largest values of the F statistics with respect to the mono-exponential fits and the largest amplitudes of the faster decay components, indicate a fast HN exchange (Tables 5 and 6).
Table 5 Averaged values of the statistics F for the comparison of bi-exponential and mono-exponential fit in two groups: nine fast HN-exchanging residues (L8-T12, A46, and L73-G75) and the remaining sixty one. The value of the statistics F for a probability level 0.01, F = 10.9, is given as referenceTable 6 Fractions, f = Afast/(Afast+Aslow), of the fast amplitudes of the exponential decay determined in bi-exponential fits of R1 data. Fractions in the table are averaged over the fast HN exchanging residues (L8-T12, A46, and L73-G75) and the remaining sixty-oneAn example of a bi-exponential fit to experimental data for fast exchangeable HN residue A46 is shown in Fig. 10. The deviation of the mono-exponential R1 value from the slow component in bi-exponential fit is equal to 6.7% and 0.3% for H2O/D2O and H2O solutions, respectively. For less demanding applications, the mono-exponential fit for H2O solution data is sufficiently accurate, and its variation from the bi-exponential fit is comparable to the accuracy of the measurement. In many cases, this allows a simplification of experimental determination of R1. Such solvent-induced effect is independent of magnetic field strength, detection method (HSQC, TROSY), or interference suppression method (selective or nonselective HN pulses), (Supplementary Material). As before, a key element in such a distinction is the presence of D2O.
Fig. 10
R1 data for A46 residue measured in different solvent (with 10% D2O - upper panel and without D2O - lower panel) at 22.3 T using HSQC method and HN band selective irradiation. Parameters determined in bi-exponential fit for upper panel: F = 278, fast component (Af=181, R1f = 10.8 s− 1), slow component (As=1033, R1s = 1.399 ± 0.009 s− 1), mono-exponential fit (A = 1098, R1 = 1.493 ± 0.030 s− 1). Parameters determined for lower panel: F = 40, fast component (Af=34, R1f = 13.8 s− 1) slow component (As=1110, R1s = 1.446 ± 0.003 s− 1), mono-exponential fit (A = 1114, R1 = 1.451 ± 0.002 s− 1)
In summary, R1 measurements made in the 100% H2O using HN-band selective pulses for interference suppression appear to be the least susceptible to experimental artifacts.
Comments (0)