
Remember me
To identify relevant LHb-projecting afferents, we injected fluorogold (FG) into the medial LHb (Fig. 1A, B). In situ hybridization of CB1R and glutamic acid decarboxylase (GADs, isoforms 65 and 67) transcripts was performed for co-localization in FG immunoreactive (FG-IR) cells projecting to the LHb. Coronal brain sections through the basal forebrain (BF) corresponding to caudal NAcs, and rostral VP were obtained from 3 rats over a span of ~2.0 mm (+2.0 mm to +0.0 mm) rostral to bregma [22]. Moderate FG-IR was found in NAcs beginning from ~+1.4 mm to bregma and extending to ~+0.7 mm (Fig. S1, Supplemental Materials). Lighter FG-IR was also localized to cells in the horizontal and vertical limbs of the diagonal band of Broca (DB, Fig. S1), and some of these DB cells also expressed the cholinergic marker, choline acetyl transferase (ChAT, not shown). FG-IR labeling of cells in NAcs and VP was also noted in more caudal sections, ~0.6–0.9 mm anterior to bregma (Fig. S1). Thus, FG-IR was densest in more caudal regions of the NAcs and the most rostral region of the VP, and some localization was found in the DB. Co-localization of CB1R and GADs mRNAs was observed in 38% of the FG-IR cells in VP (Fig. 1D), and in 18% of the FG-IR NAcs neurons (Fig. 1C–E). These findings are generally consistent with studies describing FG-IR localization after LHb injections in mice and rats [23,24,25,26].
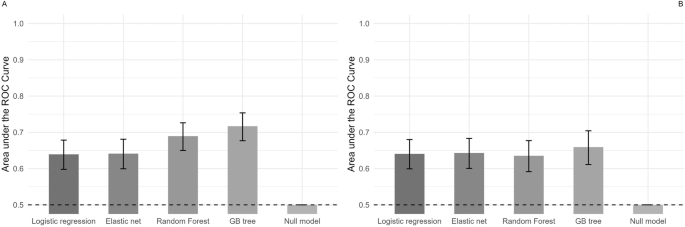
Fig. 1: Basal forebrain (BF) neurons located in the nucleus accumbens shell (NAcs) and the ventral pallidum (VP) project to the LHb and express CB1R mRNA, GADs mRNA, or both.
A Retrograde tracer FG was delivered into the LHb. B FG-IR in the injection site (brown). C Low magnification of a NAc shell (d) and VP (e) coronal section showing detection of FG-immunoreactivity (FG-IR, blue), expression of CB1 mRNA (red), and expression of GADs mRNA (green). These areas in yellow boxes are shown at higher magnification in the sequence shown in (E), top and bottom, respectively. D Number of FG-IR cells detected in VP and NAcs, sorted by co-expression of CB1R mRNA and/or GADs mRNA. A total of 208 FG-IR VP neurons projecting to the LHb were detected, and of these 24 (13%) expressed only CB1R mRNA, 59 (28%) expressed only GADs mRNA, 80 (38%) co-expressed CB1R and GADs mRNA, and 45 (21%) lacked both CB1R and GADs mRNA. A total of 369 FG-IR NAcs neurons projecting to the LHb were detected, and of these 97 (26%) expressed only CB1 mRNA, 79 (21%) expressed only GADs mRNA, 67 (18%) co-expressed CB1R and GADs mRNA, and 126 (34%) lacked both CB1R and GADs mRNA. FG-IR cell counts were made between +1.92 and +0.72 mm from bregma (n = 3 rats, 8–9 sections per rat). E NAcs and VP areas corresponding to boxes (d, e) in panel (C) at higher magnification. FG-IR neurons expressing CB1R mRNA without GADs mRNA are indicated by single arrows, FG-IR neurons expressing GADs mRNA and CB1R mRNA are indicated by single arrow heads, and double arrow heads indicate FG-IR without GADs or CB1R mRNA in NAcs. A FG-IR neuron expressing GADs mRNAs, without CB1R mRNA in VP is indicated by double arrows. Abbreviations; aca, anterior commissure; LHb, lateral habenula; MHb, medial habenula; fr, fasciculus retroflexus.
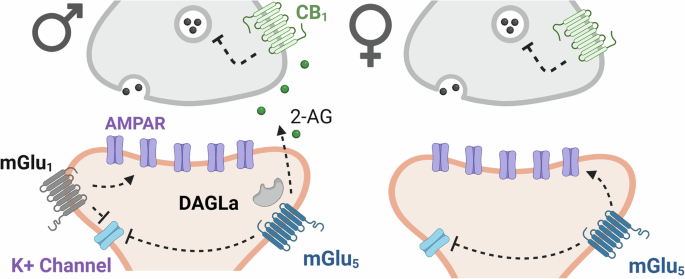
Synaptic inputs from BF to LHb are primarily GABAergicTo assess the influence of afferents on LHb neurons in wildtype rats, AAV5-hSyn-ChR2-eYFP was injected into VP/NAcs areas where retrograde FG labeling was observed, or for comparison, into the interfascicular nucleus of the VTA, a mixed GABAergic/glutamatergic LHb input (Fig. 2A, B) [27]. Electrophysiology was conducted in LHb neurons ~8 weeks after transfection. Photostimulation of axon terminals in LHb arising from VTA or VP/NAcs neurons evoked synaptic currents in LHb neurons (Vhold = 0 mV) (Fig. 2C, D). The synaptic currents at both inputs were eliminated by the GABAA receptor channel blocker picrotoxin (PTX, Fig. 2C–F). The photoactivated optical IPSCs (oIPSCs) from both projections were also eliminated by the Na+ channel blocker, tetrodotoxin (TTX), and this was reversed by the Kv1 K+ channel blocker 4-aminopyridine (4-AP) [27], confirming that these currents are monosynaptic (Fig. 2C–F). Additionally, time constants for the rise (Fig. 2G) and decay (Fig. 2H) kinetics of the currents were similar at both pathways (unpaired t test; t13 = 0.79, p = 0.45, and t13 = 0.59, p = 0.73, respectively). Additionally, the peak synaptic currents evoked at 0 mV membrane potential were not significantly affected by AMPAR (DNQX) or glycine receptor (strychnine) antagonists (Fig. 2I; mixed effects, repeated measures ANOVA, main effect of time, F2.407, 33.42 = 3.07, p = 0.0512). Similarly, the decay time constants for these currents were unaffected by DNQX or strychnine (Fig. S2), suggesting that glutamate and glycine receptors did not make large contributions to the synaptic currents under these conditions (also see [19]). Membrane voltage versus synaptic current amplitude plots indicated reversal potentials near that predicted for Cl- (ECl) under our recording conditions (Fig. 2J). Thus, considering a calculated liquid junction potential of −9.2 mV, we predicted ECl = −61.22 mV, and the mean measured ECl values were: VTA→LHb, ECl = −56.82 mV, 95% CI = −61.14 to −52.93 mV; NAcs→LHb mean ECl = −56.47 mV, 95% CI = −59.41 to −53.72 mV; (Fig. 2J). Together, these data show that photoactivation of BF or VTA afferents inhibits LHb neurons via activation of GABAA receptor/Cl- channels.
Fig. 2: Synaptic currents evoked by ChR2 stimulation in LHb after transfection of VP/NAcs or VTA in are primarily mediated by GABA and not glutamate or glycine.
A Diagram showing sites of ChR2 construct injection into VP/NAcs and photomicrographs showing eYFP fluorescence at the injection site and in the habenula ~8 weeks after injection. B Diagram showing injection sites and eYFP after VTA injections of ChR2 construct. Number at left indicate sections relative to bregma. Mean photostimulation-evoked current sweeps obtained during control periods, and during sequential application of TTX, 4-AP, and PTX in LHb neurons from rats transfected with AAV-ChR2 in the VP/NAcs (C) or VTA (D). The representative mean time courses for these experiments are shown in (E, F). In (G), the 10–90% rise times are shown for both inputs to LHb, and (H) shows the time constant for the decay of the synaptic currents (tau) evoked by each LHb input. These kinetic measures were similar for both pathways (unpaired t test; rise time, t13 = 0.79, p = 0.45, decay time constant, t13 = 0.59, p = 0.73, respectively). I Mean time course for synaptic currents evoked via photostimulation at each pathway during application of the AMPAR antagonist DNQX (10 µM) and glycine receptor antagonist strychnine (5 µM) at a holding potential of 0 mV. J Mean current-voltage (I-V) relationships for photoactivated synaptic currents from VP/NAcs and VTA inputs to LHb neurons. The calculated reversal potential for Cl- is indicated by downward arrow. Above are signal averaged synaptic currents collected during activation of each pathway across a range of membrane holding potentials (Vm). The synaptic current I-V curves reversed near that predicted for Cl- ions (ECl-, see text). Number of cells/rats: (E), 5/3; (F), 6/4; (G–I), 10/16; (J), VTA, 2/3; VP/NAcs, 4/7.
Inhibitory control of LHb neuron excitability by VP/NAcs and VTA inputs to LHb in wildtype ratsTo determine the influence of these inputs on LHb neuron excitability, we performed whole-cell recordings in brain slices from rats injected with AAV-ChR2 in VP/NAcs or VTA. Plotting the relationship between photoactivated, oIPSC amplitudes and laser intensity, we found that the VP/NAcs inputs to LHb generated significantly larger oIPSCs than those from VTA afferents (2-way RM ANOVA Interaction F5,90 = 8.19, p < 0.0001; pathway main effect F1,18 = 8.231, p = 0.01, Fig. 3A).
Fig. 3: Relative strength of inhibition of LHb neurons by VTA and VP/NAcs afferents and sensitivity to cannabinoids in vitro.
A Relationship between 473 nm laser power (single -pulses, 2 ms duration) and oIPSC amplitude in LHb neurons from wildtype rats injected with AAV-ChR2 in either VTA or VP/NAcs. VP/NAcs inputs to LHb generate significantly larger oIPSCs than those from VTA. B, C Action potentials (AP) generated by injection of +200 pA (1 s) currents in LHb neurons. Waveforms in gray were recorded without activation of VP/NAcs or VTA inputs, whereas AP wave forms shown in blue were recorded during stimulation of ChR2 by 473 nm light (blue circles) at VP/NAcs (B) or VTA (C) inputs (1 s photostimulation train = seven-2 msec duration pulses delivered at 7 Hz at 145 ms intervals during LHb neuron depolarization). D Summary of effect of activation of VP/NAcs (n = 18) or VTA (n = 24) LHb input by ChR2 on the probability of AP discharge. Activation of either input significantly reduced AP probability, but the VP/NAcs input was significantly more effective at silencing LHb neurons t40 = 3.526, p = 0.011, (unpaired t test). E Effect of bath application of the cannabinoid agonist WIN55212-2 (WIN, 2 µM) on oIPSCs evoked by ChR2 at VP/NAcs (n = 18 cells) or VTA (n = 8 cells) inputs to LHb neurons. Whereas WIN had a small effect on IPSCs from VTA input, the effect on VP/NAcs input was significantly larger (p = 0.0001, t test). The Y-axis title for bar graph is the same as time course. (F). WIN significantly increased the paired-oIPSC ratio (PPR) at VP/NAcs inputs to LHb, indicating a presynaptic effect (paired t test, t7 = 4.7, p = 0.022). G Preincubation of LHb cells with the neutral CB1R antagonist NESS 0327 (NESS) prevents the inhibition of VP/NAcs oIPSCs in LHb (one-sample t test, WIN = t7 = 6.76, p = 0.0003; WIN + NESS = t7 = 1.16, p = 0.284). Left and right graph share y-axis. H CB1R antagonism by NESS reveals tonic eCB suppression of VP/NAcs oIPSCs in LHb neurons. Left and right panels share y-axis label. I AP waveforms generated by current injection with ChR2-activation of VP/NAcs inputs (red) or without ChR2-activation (gray) in control aCSF. J AP waveforms recorded with and without ChR2-activation of VP/NAcs inputs to LHb during WIN application. K AP probability before (gray circles) and during application of WIN (red circles). The reduction of synaptic inhibition by WIN significantly increased AP probability (paired t test, t9 = 4.76, p = 0.001). Number of cells/rats: A VP/NAcs to LHb: 8/5, VTA to LHb, 8/5; (D) VP/NAcs to LHb, 18/13, VTA to LHb, 24/16; (E) VP/NAcs, 8/6, VTA, 8/5; (F). 8/5; (G). WIN alone, 8/5: WIN + NESS, 8/5; (H) 11/8; (K) 10/7.
To examine how these inhibitory inputs control LHb neuron excitability, ChR2 expressed on these axons was photoactivated using brief pulses of light (7 pulses, 145 ms interval), coincidentally with LHb neuron membrane depolarization (1 s, 200 pA) sufficient to generate action potentials (AP) in current clamp (Fig. 3B, C). The probability of AP firing was defined as the number of APs generated by depolarization in the absence of photostimulation, divided by that recorded during photoactivation of the afferents. Although, photostimulation of both LHb inputs reduced AP probability, this was significantly larger for the VP/NAcs BF input compared to that from VTA axons (Fig. 3D, t40 = 3.526, p = 0.011, unpaired t test). Thus, whereas both VP/NAcs and VTA afferents can inhibit LHb neuron excitability, the VP/NAcs input can more effectively silence these glutamate neurons.
CB1Rs inhibit VP/NAcs input to LHb neurons to control excitability and are tonically activated by eCBsOur FG-IR experiments showed co-localization of CB1R mRNA in BF cells from VP/NAcs projecting to the LHb (Fig. 1). Therefore, we measured CB1R activation effects on VP/NAcs and VTA synaptic input to LHb. Photostimulated oIPSCs from VP/NAcs inputs were significantly inhibited by the CB1R agonist WIN55,212-2 (WIN, 2 µM; mean inhibition = 65.3% of control, 95% CI = 56.48–74.13% control; one sample t test, VP/NAcs = t17 = 8.30, p < 0.0001). The oIPSCs evoked by photostimulation of VTA inputs were also slightly inhibited by WIN (mean inhibition = 94.56% of control, 95% CI = 91.51 to 97.62% control; one sample t test, VTA= t7 = 4.30, p = 0.004). However, CB1R inhibition of oIPSCs elicited from the VP/NAcs inputs was significantly greater than that at VTA afferents (unpaired t test = t24 = 4.57, p = 0.0001, mean difference = 29.26% control, 95% CI = 16.05–42.47% control, Fig. 3E).
To determine whether the inhibition of oIPSCs by WIN occurred through a presynaptic mechanism on VP/NAcs axons, we measured paired oIPSC responses, evoked by 2 rapid photoactivations of ChR2 (100 ms interval), and calculated the ratio of first to the second oIPSC (paired-pulse ratio, PPR = oIPSC2/oIPSC1). The mean PPR was less than unity before WIN application, suggesting inhibition of the second oIPSC relative to the first, and there was a significant increase in the PPR during WIN application (two-tailed t test, t7 = 4.7, p = 0.022 Fig. 3F). This indicates that WIN decreased GABA release probability from VP/NAcs axons in the LHb. To confirm involvement of CB1Rs, we tested the effect of WIN in brain slices pre-incubated in the neutral CB1R antagonist, NESS0327 (NESS, 500 nM [28]). We found that NESS blocked the inhibition of oIPSCs by WIN (Fig. 3G, WIN alone, mean and 95% CI; 70.05% control, and 60.42–81.47% control; WIN + NESS, 97.57% control 83.37 to 98.52% control; one-sample t test, WIN = t7 = 6.76, p = 0.0003; WIN + NESS = t7 = 1.16, p = 0.284). This indicates that WIN inhibited VP/NAcs oIPSCs in the LHb by activation of CB1Rs on these axon terminals.
To determine whether endogenous cannabinoids (eCB) might also activate CB1Rs on VP/NAcs axons in LHb, we examined the effect of NESS alone on these responses. We found that NESS significantly increased the size of the oIPSCs, suggesting that they were tonically inhibited by an eCB acting at CB1Rs (Fig. 3H, paired t test = t10 = 3.64, p = 0.0046, mean %control and 95% CI = 115.91% control and 106.156–125.66% control, n = 11 neurons).
To determine the consequence of CB1R inhibition of GABA release from VP/NAcs axons on LHb neuron excitability, we next measured the effect of photostimulation of these axons on AP probability before, and during application of WIN. As described above (Fig. 3B, D), ChR2-activation of VP/NAcs inputs strongly inhibited depolarization-induced APs in LHb neurons before WIN was applied (Fig. 3I; mean (±SEM) AP probability = 0.158 ± 0.083, Fig. 3K). However, AP firing probability was significantly increased to 0.479 ± 0.060 during WIN application (Fig. 3J, K, paired t test, t9 = 4.76, p = 0.001, mean difference = 0.3115, 95% CI of difference = 0.1633–0.4597, n = 10 neurons). Together, these data show that CB1Rs strongly control GABA release from VP/NAcs afferents to LHb neurons, and that activation of these receptors increases LHb neuron excitatory output.
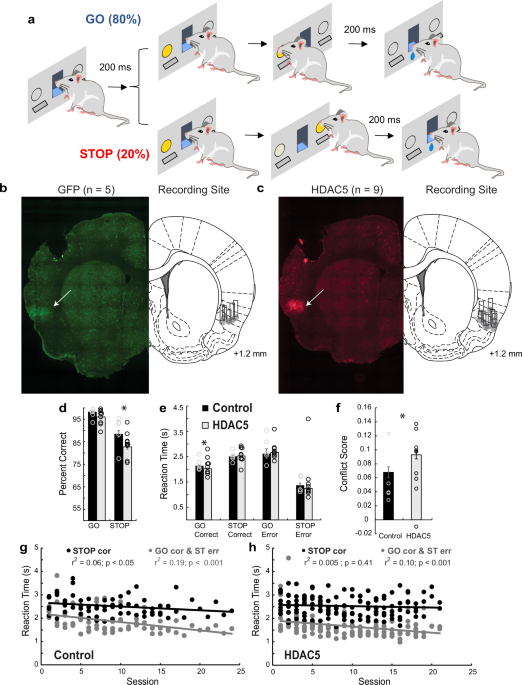
LHb Involvement in response inhibition in a food reinforced operant taskThe electrophysiology experiments indicated that BF inputs to LHb strongly inhibit LHb activity, and this inhibition is decreased by CB1Rs. To examine a potential role for this afferent in LHb-involved impulsive behavior, we used a food-reinforced discrimination task that avoided potentially disruptive psychomotor effects of cocaine as a reinforcer [13]. Here, male and female wildtype rats were trained in a DS-NS task using a specific auditory tone (DS) to signal reward availability for responses on an active operant lever. Conversely, white noise, served as a neutral stimulus (NS), indicating no reward for lever presses. Training proceeded until criteria of active lever responding in ≥90% of the DS trials, and ≤30% of NS trials was achieved. To validate LHb involvement in this task, we next examined performance on DS and NS trials during pharmacological and neurobiological manipulations that decreased response inhibition in the 5CSRTT and cocaine-rewarded Go/NoGo paradigm [13,14,15]. Infusion of scopolamine into LHb significantly decreased the percentage of DS trials in which responding was observed, whereas infusion of B/M did not significantly alter this measure (Fig. 4A). However, these pharmacological manipulations did not alter the number of responses on the inactive operant lever (Fig. S3A) or the number of food port entries (Fig. S3B). Importantly, like our previous studies, B/M, or scopolamine infusion into LHb significantly increased NS trials in which responding was observed, whether analyzed as percent of NS trials, or total responses during NS presentation (Fig. 4B, C).
Fig. 4: Effects of pharmacological manipulation of the LHb on response inhibition in wildtype rats.
A–C Effects of intra-LHb infusion of Saline (Sal), baclofen/muscimol (B/M) or scopolamine (Scop) on DS-NS responding (n = 8 male and 3 female rats). A Scop infusion into LHb significantly decreased responding during trials in which food pellet availability was signaled (DS, repeated measures 1-way ANOVA, F1.64,16.4 = 6.95, p = 0.009, p values from Dunnett’s post hoc test). B Scop and B/M significantly increased the percentage of NS trials (when reward was not available) in which responses occurred (repeated measures 1-way ANOVA, F1.4,14 = 7.79, p = 0.009, p values from Dunnett’s post hoc test). C Scop and B/M significantly increased the number of NS trial responses (repeated measures 1-way ANOVA, F1.67,16.73 = 3.87, p = 0.0478, p values from Dunnett’s post hoc test). D–G Effects of systemic injection of Δ9-THC (1 mg/kg) or vehicle on DS-NS responding in male and female rats. D Δ9-THC injection increased the percentage of NS trials in which responses were observed in male rats only (2-way mixed effects ANOVA, n = 8 males and 6 females, drug x sex interaction, F1,10 = 12.57, p = 0.0053). The numbers above bars in (D–G) represent post hoc comparison p values using the uncorrected Fisher’s Least Significant Difference test (uFLSD). The legend in (D) applies to panels (D–F). E Systemic Δ9-THC effect on the number of responses during NS trials in male and female rats (2-way ANOVA, Drug x Sex Interaction, F1,24 = 6.50, p = 0.0165). F No effect of Δ9-THC on percent of trials responding when reward availability was signaled (DS; 2-way mixed effects ANOVA, n = 10, F1,18 = 3.54, p = 0.080). G The increase in proportion of NS trial responses caused by systemic injection of Δ9-THC in male rats was prevented by infusion of AM251 into the LHb (n = 10; 2-Way RM ANOVA, systemic THC x AM251 infusion Interaction = F1,18 = 9.35, p = 0.0068).
In a previous study, we also found that response inhibition in the 5CSRTT and Go/NoGo tasks was impaired by systemic injection of Δ9-THC (1 mg/kg) [15], and our present study and others show that some LHb afferents express CB1Rs [18, 19, 29, 30] (Fig. 1). Therefore, we examined the effect of Δ9-THC on DS-NS behavior and found that response inhibition was significantly impaired by systemic injection of the phytocannabinoid, although this was observed in only male rats (mixed effects ANOVA, drug x sex interaction F1,10 = 12.57, p = 0.0053, Fig. 4D, E). In addition, Δ9-THC did not alter the number DS trials in which responding was observed in males or females (F1,18 = 3.454, p = 0.0795, mixed effects ANOVA, Fig. 4F). However, we did observe a significantly higher number of inactive lever responses in males compared to females (Fig. S3C), but this measure was not significantly altered by systemic Δ9-THC (Fig. S3C). Our prior studies also showed that infusion of the CB1R antagonist AM251 into LHb prevented disruption of response inhibition by systemic Δ9-THC in the 5CSRTT and Go/NoGo paradigms [15]. Similarly, intra-LHb AM251 also blocked the pro-impulsive effects of systemic Δ9-THC in the present DS-NS task in male rats (Fig. 4G, n = 10; 2-Way RM ANOVA, Interaction = F1,18 = 9.35, p = 0.0068, p = 0.0005, Uncorrected Fisher’s Least Significant Difference (uFLSD) post hoc). Therefore, similar to our previous measures of impulsivity, response inhibition in the DS-NS paradigm requires intact LHb function and is impaired by disruption of LHb cholinergic signaling or by systemic injection of Δ9-THC.
VP/NAcs projections to LHb influence response inhibition in the DS-NS task in GADCre ratsTo determine whether the GABAergic inputs to LHb arising from the VP/NAcs influence response inhibition, we trained GADCre rats [20] in the DS-NS paradigm, 7–8 weeks after infusion of viruses expressing the inhibitory halorhodopsin (AAV- EF1α-DIO-eNpHR3.0-eYFP (NpHR)), excitatory Channelrhodopsin-2 (AAV5-hSyn-ChR2-eYFP, ChR2), or enhanced yellow fluorescent protein alone (AAV-Control; AAV5- hSyn1-eYFP) into the VP/NAcs. We then assessed the effects of photostimulation of VP/NAcs axon terminals in the LHb (NpHR, 545 nm, constant pulse, alternating 5 s on/off; ChR2 and eYFP, 455 nm, 60 Hz pulses, alternating 5 s on/off) on operant responding during DS and NS trials. Photostimulation of the LHb failed to alter NS responding in rats receiving eYFP (Fig. 5A) or NpHR constructs into the BF (Fig. 5B). However, photoactivation of ChR2 in the LHb significantly increased the percentage of NS trials in which responses were observed (Fig. 5C, D), as well as the total number of these responses (Fig. S4A) in GADCre rats. This impairment in response inhibition caused by photostimulation of ChR2 on this pathway did not significantly differ between male and female rats (Fig. 5D; n = 16 male and 11 female rats; 2-way RM ANOVA, light x sex interaction, F1,25 = 1.546, p = 0.225; main effect of light, F1,25 = 41.03, p < 0.0001; Post hoc comparisons by uFLSD, effect of light in males, p < 0.0001; effect of light in females, p = 0.0025). In contrast, photostimulation of ChR2 in LHb did not significantly affect responding during DS trials (Fig. S4B), the number of food port entries (Fig. S4C), nor the number of responses on inactive levers (Fig. S4D). Similar results were observed in optogenetic experiments in wildtype rats (Fig. S5). The data suggest that increased synaptic inhibition of LHb neurons via stimulation of VP/NAcs GABAergic axons increases impulsivity, as measured by decreased response inhibition in the DS-NS task.
Fig. 5: Impaired response inhibition during activation of GABAergic BF input to the LHb by ChR2 in GADCre rats.
A Comparison of the percent of neutral stimulus (NS) trials in which responses on the active lever were observed in the absence (off) and presence of 455 nm light delivered via optical fibers terminating above the LHb, ~8 weeks after expression of Cre-dependent eYFP in the VP/NAcs (n = 6 rats; paired t test, t5 = 1.67, p = 0.155). B Optogenetic inhibition of VP/NAcs GABAergic inputs to LHb by stimulation of NpHR with 545 nm light does not affect NS responding (n = 4 rats; paired t test, t3 = 0.38, p = 0.728). C Activation of VP/NAcs GABAergic inputs by light stimulation of ChR2 in LHb significantly increased the percent of NS trials in which responses were observed (paired t test; n = 10 male rats, t9 = 3.44, p = 0.007). The total number of NS responses shown in Fig. S4A. D ChR2 photostimulation with 455 nm light significantly increases responding during presentation of the NS, and this is not significantly different between male and female GADCre rats (n = 16 male and 11 female rats; 2-way RM ANOVA, light x sex interaction, F1,25 = 1.546, p = 0.225; main effect of light, F1,25 = 41.03, p < 0.0001; Post hoc comparisons by uFLSD, effect of light in males, p < 0.0001; effect of light in females, p = 0.0025). ns, non-significant.
Comments (0)