
Remember me





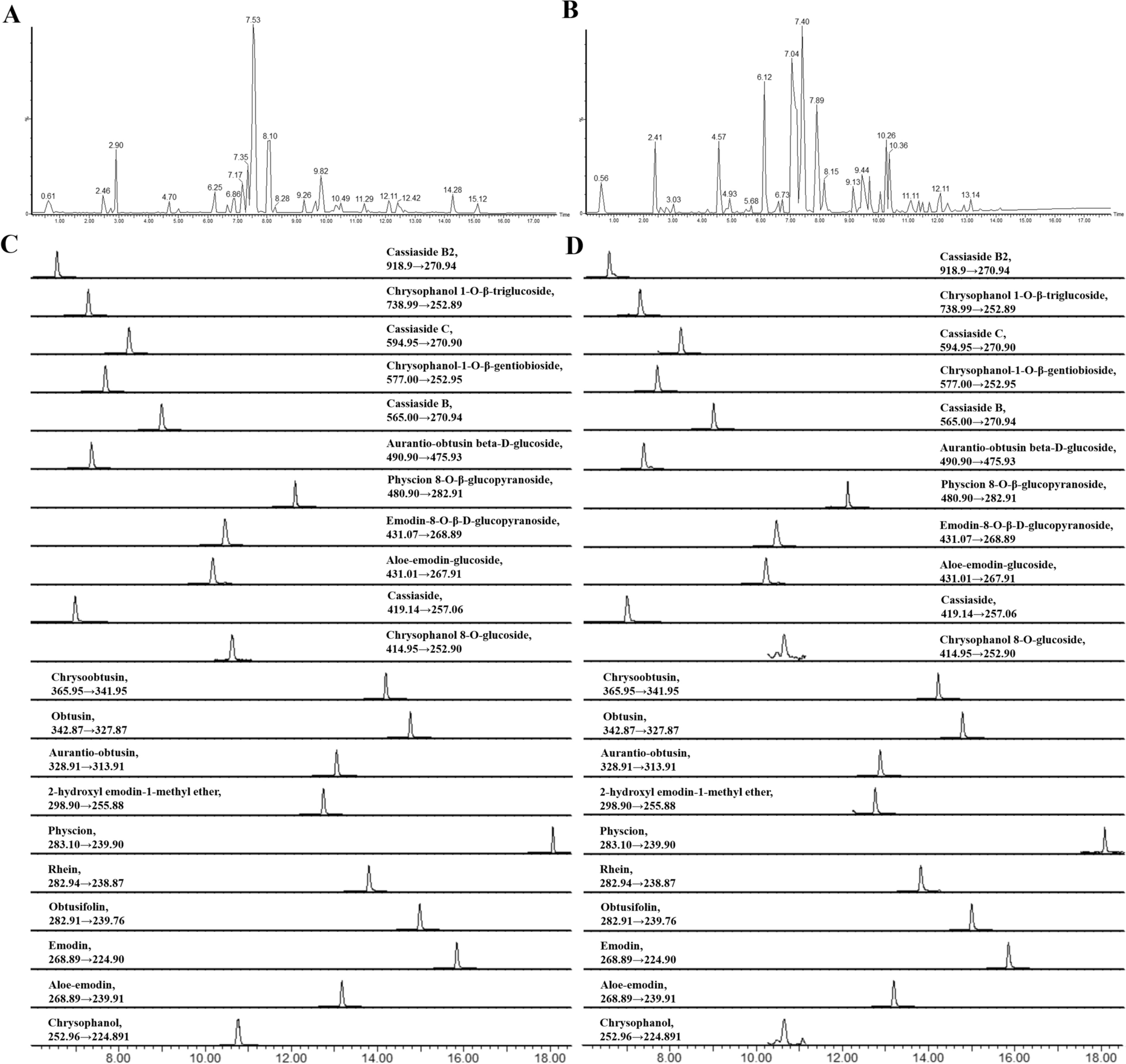

A key step in understanding the pathogenesis of liver cancer and developing intervention strategies is the construction and analysis of weighted gene regulatory networks. The PPI network was made up of public PPI databases, including BioGRID, STRING, Dip, HPRD, Mint and Intact. To construct the weighted gene regulatory network for liver cirrhosis, 897 pathogenic genes associated with liver cirrhosis were obtained from GeneCards and DisGeNET (Additional file 2: Table S2) and mapped to the PPI network. The weighted gene regulatory network consisted of 744 nodes and 4201 edges (Fig. 2). Among the top 30 genes in the gene regulatory network, 7 genes are enriched in alcoholic liver disease (hsa04936), including TNF, IL6, IFNA1, CYP2E1, ALDH2, ADH1B, ADH1C. Alcoholic liver disease usually develops fatty liver initially and can develop into alcoholic hepatitis, hepatic fibrosis and cirrhosis. The result indicated that the weighted gene regulatory network together with the weighted gene are capable of reflecting the underlying pathogenesis of liver cirrhosis. This information serves as a dependable reference for the subsequent construction of the functional response space.
Fig. 2
Weighted gene regulatory network of cirrhosis. Node size represents the relevance score of pathogenetic genes multiplied the number of literatures of liver cirrhosis, followed by extracting the square root. The red nodes indicate the top 16 pathogenetic genes with the highest weight in this network
Collection of chemical components and active components screeningChinese medicinal formulation contains multiple components, but not all of components are satisfied with pharmacokinetic screening characteristics. According to the Lipinski five rules, 267 active components were selected out from 386 components of WLP (Additional file 3: Table S3). Among 267 active components, GZ includes 194 active components, BZ includes 43 active components, ZX includes 23 active components, FL includes 7 active components and ZL includes 9 active components (Table 1). The chemical composition analysis provides a reference for selecting the active components of WLP to further analyze. The components with high concentration in the prescription usually have certain chemical activity, so we collected the chemical components with high concentration verified by experiments from the literature. The concentration of 25 components in the chromatograms of WLP were found (Table 2).
Table 1 WLP components collected in the public database and active components after selectingTable 2 Collection and concentration arrangement of chemical componentsShared components of herbs in WLPThere are 9 shared components of two herbal medicines in WLP (Fig. 3). For example, caprylic acid (WLP1) is a common component in two herbal medicines (GZ, FL). Caprylic acid is a medium-chain fatty acid (MCFAs). The in vitro experiments with purified BCKDC-BDK complex showed that caprylic acid inhibited BDK activity. Rats were orally administrated with caprylic acid to down-regulate BDK activity by reducing the number of BDK binding complexes, thereby activating liver BC-KDC, decreasing plasma BCAA concentration and increasing serum ketone body concentration [35]. Hemo-sol (D-limonene, WLP7) is a common component of two herbal medicines (GZ and BZ). It has been found to be effective in preventing carbon tetrachloride-induced liver fibrosis in rats by controlling oxidative stress and suppressing the inflammatory pathway. Furthermore, it can regulate the expression of markers associated with fibrosis, such as TGF-b and hydroxyproline [36].
Fig. 3
The UpSet picture of WLP components
Specific components of herbs in WLPMost herbs have their own specific components in addition to the common ones. L-lysin (WLP207), a specific component of BZ, is able to regulate the increase in B-oxidation cells in the liver of SAMP8 mice to which the L-lysin-rich feed was added, suggesting that the dietary intake of lysine could prevent liver steatosis by stimulating B-oxidation in SAMP8 mice [37]. Specific component NON (Decanoic acid, WLP116) of GZ could be converted into decanoate, which upregulates the expression of adipogenic transcription factors and enzymes to reduce fat formation [38]. HCI (3-phenylpropionic acid, WLP163), which is a particular component of GZ, can reduce the CISP-induced oxidative stress in liver. This is achieved by limiting inflammatory signals like COX-2 and NF-κB, as well as apoptotic signals including BAX and caspase-3. Moreover, it enhances the expression of Bcl-2. [39].
C-T Network construction of active componentsThe treatment of complex diseases with TCM formulas involves the chemical components and their targets. A single component can correspond to multiple targets, and likewise, a single target can be targeted by multiple components. Therefore, it can be viewed as a network of multiple components and multiple targets, known as the C-T network. To study the mechanics of WLP in addressing liver cirrhosis, the C-T network was established using 267 active components along with 1372 targets. Due to the potential for multiple targets corresponding to a single component, there is a total of 10,849 relationship pairs existing between the mentioned components and their respective targets. The mean number of targets per component was 40.63, indicating the multi-target properties of WLP in treating liver cirrhosis. Caprylic acid (WLP1, degree = 252) has the most targets, with Benzyl acetate (WLP65, degree = 248), WLN: 2OVR (formic acid, WLP30, degree = 244), HCI (3-phenylpropionic acid, WLP163, degree = 252), BZM (benzoic acid, WLP83, degree = 220), and PHA (phenylalanine, WLP218, degree = 214) following closely behind.
The average target degree of the components is 7.91 in the C-T network. The top 20 by weight are MAPT, CA3, CES1, CES2 and MOAB, etc. These targets have been reported to be related to liver diseases and may play a crucial role in the therapeutic effects of WLP in treating liver cirrhosis. Research has indicated that MAPT is able to predict the prognosis of patients with liver cancer [40]. CA3 can inhibit the proliferation of hepatoma HepG2 cells and induce their apoptosis [41]. For immune liver injury, the expression and metabolic activity of CES1 and CES2 are significantly decreased [42]. MAOB is involved in the biosynthesis of endogenous geranyl valeric acid (GGA) through geranyl geraniol oxidation, while GGA is a prophylactic agent for secondary primary liver cancer [43]. Overall, these results indicate that WLP can treat liver cirrhosis via multiple targets and confirm its multi-targeted role in treatment.
Determination and verification of function response spaceWe established a multi-objective optimization model including three customized objective functions, to identify the subset with the largest degree from the C-T network as the FRS. The FRS consists of 993 nodes and 2493 edges. 940 nodes from the targets and 53 nodes from the components, thus we defined 940 effective proteins from the FRS.
To determine whether the effective proteins identified from the FRS adequately representing the pathogenic genes of cirrhosis at a functional level, we conducted functional pathway analyses using effective proteins, components, and pathogenic genes specific to cirrhosis. Notably, the shared genes of targets and pathogenic genes (CAP) were enriched in 108 KEGG pathways (P < 0.05), while effective proteins were enriched in 176 KEGG pathways (P < 0.05), with effective proteins’ enrichment pathways expected to encompass 83% of the pathways enriched by CAP (Fig. 4A). Additionally, KEGG pathway analyses were performed on disease-specific targets and component-specific targets to assess whether the FRS could be substituted by these targets. Disease-specific targets covered 56% of the pathways enriched by CAP, and component-specific targets covered 69% of these pathways (Figs. 4B and C), which were markedly lower than the coverage of effective proteins. Furthermore, effective proteins’ pathways were largely included in both component target pathways and disease target pathways (Fig. 4D). These findings validate the accuracy and robustness of our multi-objective optimization and establish that the effective proteins identified in the FRS play a crucial role in the pathogenesis of cirrhosis.
Fig. 4
Verification of Function Response Space. A Venn diagram shows the number of pathways enriched by effective proteins and the shared genes of CAP. B Venn diagram shows the number of pathways enriched by component-specific targets, disease-specific targets and CAP. C Proportion histograms depict the enriched pathways of component-specific targets, disease-specific targets and effective proteins in FRS and CAP. D Bubble diagram shows the shard pathway enriched by effective proteins, component targets and disease targets (liver cirrhosis pathogenic genes)
According to KEGG pathway analysis, these potent proteins were involved in non-alcoholic fatty liver disease (hsa04932), metabolism of xenobiotics by cytochrome P450 (hsa00980), drug metabolism-cytochrome P450 (hsa00982) and VEGF signaling pathway (hsa04370) (Fig. 4D). For example, non-alcoholic fatty liver disease (hsa04932) suggests that non-alcoholic fatty liver disease may further contribute to liver cirrhosis. After the formation of non-alcoholic fatty liver disease, ROS production is enhanced due to mitochondrial β-oxidation of fatty acids and oxidative stress of endoplasmic reticulum stress, leading to lipid peroxidation. Lipid peroxidation further leads to the production of cytokines (Fas ligand, TNF-α, IL-8, and TGF) and promotes cells death, inflammation, and fibrosis [44]. Polymeric immunoglobulin receptor (PIGR) is significantly increase in non-alcoholic fatty liver disease and cirrhosis. The strategy of using the multi-objective optimization model phase to optimize the herbal formulation is reliable, and the FRS may play a part in by mediating multiple metabolic and inflammation-related pathways.
KGEC selection and validationThe knapsack algorithm model was applied to optimize the active components group and obtain the KGEC. A total of 29 components were chosen as the KGEC, as they collectively contributed to 100% coverage of the targets of effective proteins (Table 3).
Table 3 The information of KGEC in WLPWe conducted KEGG pathway analysis for KGEC targets and CAP targets separately to examine the potential role of WLP in treating liver cirrhosis. 177 pathways were found to be enriched in KGEC targets (P < 0.05), while 108 pathways were enriched in CAP targets (P < 0.05). Notably, the pathways enriched in KGEC targets covered 83.33% of those enriched in CAP targets (Fig. 5A). The majority of the KEGG targets were involved in pathways such as non-alcoholic fatty liver disease (hsa04932), thermogenesis (hsa04714), cAMP signaling pathway (hsa04024), chemical carcinogenesis-reactive oxygen species (hsa05208), cytochrome P450 metabolism of xenobiotics (hsa00980), drug metabolism-cytochrome P450 (hsa00982), and oxidative phosphorylation (hsa00190) (Fig. 5B). Of these, the non-alcoholic fatty liver pathway (hsa04932) results in an increase in intracellular fatty acid-derived metabolites due to a lack of insulin receptor substrate-2 (IRS-2)-associated phosphatidylinositol 3-kinase (PI3K) activity. This damages the mitochondria of NASH patients, which in turn promotes liver fibrosis. [44]. These findings suggest that combining the FRS with the knapsack algorithm model for optimizing the herbal formulation is reliable, and the predicted KGEC may have a therapeutic effect by participating in several signal transduction and apoptosis pathways.
Fig. 5
Pathway enrichment analysis of KGEC targets. A Venn diagram for KGEC targets and CAP targets through pathway enrichment analysis. B The size of the circle expresses gene’s number enriched in the pathway, and the color expresses the significance of genes enriched in the pathway
Cellular experimental verification of KGECSimple random sampling is a form of probability sampling. The probability that each sample drawn is equal, and each unit of the sample is completely independent, thus can exclude the correlation and exclusion between samples [45]. Based on this strategy, we selected scopoletin, caryophyllene oxide and hydroxyzinamic acid in KEGC for in vitro experimental validation. Here, we used TGF-β1-induced LX2 cells as a model of liver fibrosis, a widely recognized and employed model. First, through CCK8 experiments, we determined that scopolamine was safe at concentrations below 100uM, while saponin and hydroxycinnamic acid were safe at concentrations below 40uM (Fig. 6A-C). Subsequently, we successfully induced the LX2 cell model of liver fibrosis, as evidenced by a significant increase in the expression of the liver fibrosis marker gene COL1A1 (Fig. 6D-F). We found that, at safe concentrations, all three components significantly suppressed the expression of COL1A1 in a concentration-dependent manner (Fig. 6D-F). These results indicate that scopolamine, saponin, and hydroxycinnamic acid can significantly inhibit TGF-β1-induced liver fibrosis in LX2 cells, without inducing significant cytotoxicity. This further demonstrates the therapeutic effect of the KEGC selected from the FRS in treating liver fibrosis.
Fig. 6
A-C Cellular experimental verification of KGEC. LX-2 cells were treated with scopoletin, caryophyllene oxide and hydroxyzinamic acid for 48 h and the cell viability was detected by CCK-8 with DMSO as control. D-F The mRNA levels of COL1A1 were detected by qPCR after LX2 cells were serum-starved for 6 h, induced with TGF-β1 for 24 h, and then treated with the three components at indicated concentrations for 48 h
GO enrichment analysis of KGEC targetsTo further investigate the combined effects of WLP, we conducted GO enrichment analysis on all the targets interacting with KGEC (Fig. 7). The findings revealed that KGEC targets were enriched in biological processes associated with mitochondrial electron transfer, including mitochondrial electron transfer, NADH to ubiquinone (GO:0006120, involving SNCA, NDUFB5, NDUFB7, NDUFB3, etc.), and mitochondrial respiratory chain complex assembly (GO:0033108, involving UQCRB, NDUFB5, NDUFB7, etc.), as well as mitochondrial ATP synthesis coupled electron transport (GO:0042775, including UQCRB, SNCA, NDUFB5, NDUFB7, etc.). Additionally, KGEC targets were enriched in cellular components related to the respiratory chain, such as mitochondrial respiratory chain complex I (GO:0005747, NDUFB5, NDUFB7, NDUFB3, NDUFB4, etc.), respiratory chain complex (GO:0098803, UQCRB, NDUFB5, NDUFB7, NDUFB3, etc.), and NADH dehydrogenase complex (GO:0030964, etc., NDUFA10, NDUFB2, etc.), while also participating in the regulation of redox-related enzyme activities, such as NADH dehydrogenase activity (GO:0003954, NDUFB5, NDUFB7, NDUFB3, NDUFB4, etc.), and oxidoreductase activity, acting on NAD(P)H, quinone or similar compound as acceptor (GO:0016655, NQO2, CBR1, NQO1, AKR1C4, etc.). It is noteworthy that accumulating evidence suggests a close association between mitochondrial dysfunction and the pathogenesis of liver cirrhosis, with oxidative stress promoting liver inflammation and fibrosis, and poorly regulated mitochondria leading to the overproduction of reactive oxygen species [46].
Fig. 7
GO enrichment analysis of KGEC targets. The colors orange, purple, and green represent biological processes, cellular components, and molecular functions, respectively. The varying colors in the bar graph indicate the significance of gene enrichment in GO
Additionally, the targets of KGEC were enriched for pathways that were associated with inflammation, including G protein-coupled receptor signaling pathway coupled to cyclic nucleotide second messenger (GO:0007187, involving OPRK1, ADRA2C, ADRB1, CCR, etc.), second messenger-mediated signal transduction (GO:0019932, with ADRA2C, ADRB1, MAPT, CCR3, etc.), adenylate cyclase-modulating G protein-coupled receptor signaling pathway (GO:0007188, including OPRK1, ADRA2C, ADRB1, CCR3, etc.), and phospholipase C-activating G protein-coupled receptor signaling pathway (GO:0007200, with ESR1, OPRK1, CX3CR1, C3AR1, etc.). It is important to note that the transition from chronic liver disease to liver cirrhosis involves inflammation, activation of liver cirrhosis cells, and simultaneous cortical extinction due to fibrosis angiogenesis, and vascular occlusion [47]. Studies have demonstrated that mitochondrial-DAMP released by damaged hepatocyte mitochondria triggers a potent inflammatory reaction, leading to the direct activation of liver cirrhosis cells and liver fibroblasts, resulting in the formation of liver scars and ultimately leading to cirrhosis [48]. Therefore, inflammation plays a crucial role in the development of cirrhosis. Overall, the GO analysis suggests that WLP may alleviate liver cirrhosis by modulating the biological process of electron transport in mitochondria and inflammation.
KEGG enrichment analysis of KGEC targetsLiver cirrhosis is now recognized as a dynamic process rather than a terminal disease. A clinical sub-classification of cirrhosis prognosis has been proposed, delineating four distinct stages with varying mortality rates [54]. Growing evidence suggests that non-alcoholic fatty liver disease pathway (hsa04932), thermogenesis (hsa04714), cAMP signaling pathway (hsa04024), and chemical carcinogenesis-reactive oxygen species (hsa05208) significantly impact the occurrence and progression of liver cirrhosis. In order to investigate the mechanism of WLP in treating liver cirrhosis at a systemic level, we developed a comprehensive signaling pathway using these four key molecular pathways (Fig. 8).
Fig. 8
Distribution of KGEC targets on integrated signaling pathways. Various colors represent different target types. Yellow represents WLP targets, orange depicts the pathogenic genes of cirrhosis, blue refers to the targets shared by WLP and the pathogenic gene of cirrhosis and purple represents the components of WLP
To determine the position of WLP targets within these pathways, the first three columns were designated as upstream positions, while the remaining columns represented downstream positions. Among these pathways, the non-alcoholic fatty liver pathway (hsa04932) emerged as one of the top pathways influenced by KGEC in WLP for treating liver cirrhosis. KGEC regulates nine upstream targets, including Rac1, Cdc42, and TNFα, as well as 12 downstream targets such as JNK1, AP1, and IL-1. Upstream targets TNFα and JNK1/2 activate c-Jun and AP1 proteins, resulting in the production of downstream inflammatory factors like IL-6 and IL-1, which are closely associated with cell death in the steatosis liver observed in non-alcoholic fatty liver disease. Cell death can further progress non-alcoholic fatty liver disease into liver cirrhosis [49]. KGEC also regulates targets involved in chemical carcinogenesis-active oxygen (hsa05208), appearing both upstream and downstream of the pathway. These targets include EGFR, PTP18, and Src. Furthermore, Fig. 8 demonstrates that KGEC can influence the activation of p38 and AP1 proteins downstream of the non-alcoholic fatty liver pathway, impacting the treatment of liver cirrhosis. Therefore, KGEC in WLP plays a pivotal role in the treatment of liver cirrhosis by modulating the chemical carcinogenesis-reactive oxygen species-non-alcoholic fatty liver pathway, which synergistically affects tumor proliferation, angiogenesis, metastasis, and the development of liver disease.
The cAMP signaling pathway (hsa04024) and thermogenesis (hsa04714) are also significant pathways targeted by WLP for the treatment of liver cirrhosis. Many of the targets influenced by WLP are located downstream of these pathways. For instance, RAS, RSK, and PRAS40 in KGEC are downstream targets of thermogenesis (hsa04714). WLP may modulate upstream targets CB1R and GRB2, leading to the activation of downstream targets PKA and RAS. This, in turn, affects a range of signal transduction-related proteins like CREB, LSD1, PPARγ, mTOR, and PRAS40, all of which are associated with liver cirrhosis. In the cAMP signaling pathway (hsa04024), JNK, PDE, and CREB are downstream targets. KGEC can activate cAMP through upstream target ANP, influencing various genes related to cell proliferation, survival, and metabolism such as PI3K and Akt [50]. This modulation impacts hepatocyte insulin resistance and the occurrence of non-alcoholic fatty liver disease. Consequently, KGEC in WLP can synergistically contribute to the development of cirrhosis by regulating the ANP-Akt key cascade, playing a significant role in the treatment of cirrhosis.
Verification the KGEC targets in vivoFurthermore, to validate the comprehensive pathway results, we assessed the impact of the three components scopoletin, caryophyllene oxide and hydroxyzinamic acid on the NF-κB, AMPK/p38, cAMP, and PI3K/AKT pathways through western blot. The results showed that, as compared to the control group, all four pathways were activated in TGF-β1-induced LX2 cells. However, treatment with scopoletin (100 uM), caryophyllene oxide (40 uM), and hydroxyzinamic acid (40 uM) led to the suppression of the expression of p-NF-κB, p-p38, p-PKA C, and p-AKT (Fig. 9). This suggests that KGEC exerts its anti-liver fibrosis effects, at least in part, by targeting these four pathways. This further validates the reliability of our new model and the accuracy of KGEC.
Fig. 9
Verification the KGEC targets in vivo The protein expression levels of NF-κB, p-NF-κB, p38, p-p38, PKA C, p-PKA C, AKT and p-AKT were determined by western blot. β-ACTIN is shown as a loading control. (Full-length blots/gels are presented in Additional file 4: Figure S1)
Comments (0)