
Remember me





We adhere to the guidelines of Lucking et al. (2021), Pem et al. (2021) and Maharachchikumbura et al. (2021) for introducing new taxa. The classifcation scheme follows Hyde et al. (2024a) unless the taxonomy has been updated recently. We followed the recommendation of Thines et al. (2020), all scientific names at all taxonomic ranks in the paper are presented in italics.
TaxonomyDothideomycetes O.E. Erikss & Winka
Botryosphaeriales C.L. Schoch, Crous & Shoemaker
Botryosphaeriaceae Theiss. & Syd.
Notes: Botryosphaeriaceae was introduced by Theissen and Sydow (1918) with Botryosphaeria as the type genus. Members of Botryosphaeriaceae are distributed worldwide as saprophytes, parasites, and endophytes on a variety of hosts in a wide range of habitats (Slippers and Wingfield 2007; Dissanayake et al. 2016; Batista et al. 2021; Garcia et al. 2021). Currently, more than 200 species in 22 genera are included in the family (Batista et al. 2021; Dissanayake et al. 2021; Zhang et al. 2021a; Wijayawardene et al. 2022b; Rathnayaka et al. 2023), with almost all species reported from terrestrial environments, and rarely reported from freshwater habitats (Cai et al. 2003a; Luo et al. 2004; Hu et al. 2013).
Botryosphaeria Ces. & De Not.
Notes: Botryosphaeria was established by Cesati and Notaris (1863), but they did not specify the type species. The current type species, B. dothidea (Moug.) Ces. & De Not., was proposed by Barr (1972) and has been widely accepted (Slippers et al. 2013; Sun et al. 2022). Members of Botryosphaeria are mainly known as plant pathogens of many commercial crops, such as apple (Tang et al. 2012), blueberry (Ru et al. 2022), grape (Úrbez-Torres et al. 2006), and peach (Wang et al. 2011). They have also been reported as saprophytes and endophytes (Crous et al. 2006; Liu et al. 2012; Dissanayake et al. 2016, 2021). Early Botryosphaeria species were described primarily based on their sexual morphology and host associations, with 302 epithets recorded in the Index Fungorum database (accessed on April 20, 2025). Zhang et al. (2021a) evaluated the important families and genera in Botryosphaeriales and accepted eight species in Botryosphaeria, based on morphological data combined with multi-loci phylogenetic analysis. A subsequent study by Zhang et al. (2021b) examined the type specimens of Botryosphaeria species and accepted ten species within Botryosphaeria based on morphological data combined with multi-gene phylogeny results. Several new species have since been described (Chu et al. 2021; Hattori et al. 2021b; Sun et al. 2022). All known Botryosphaeria have been reported from terrestrial habitats with none yet reported from freshwater habitats.
Botryosphaeria wangensis G.Q. Li & S.F. Chen, Persoonia 40: 84 (2017)
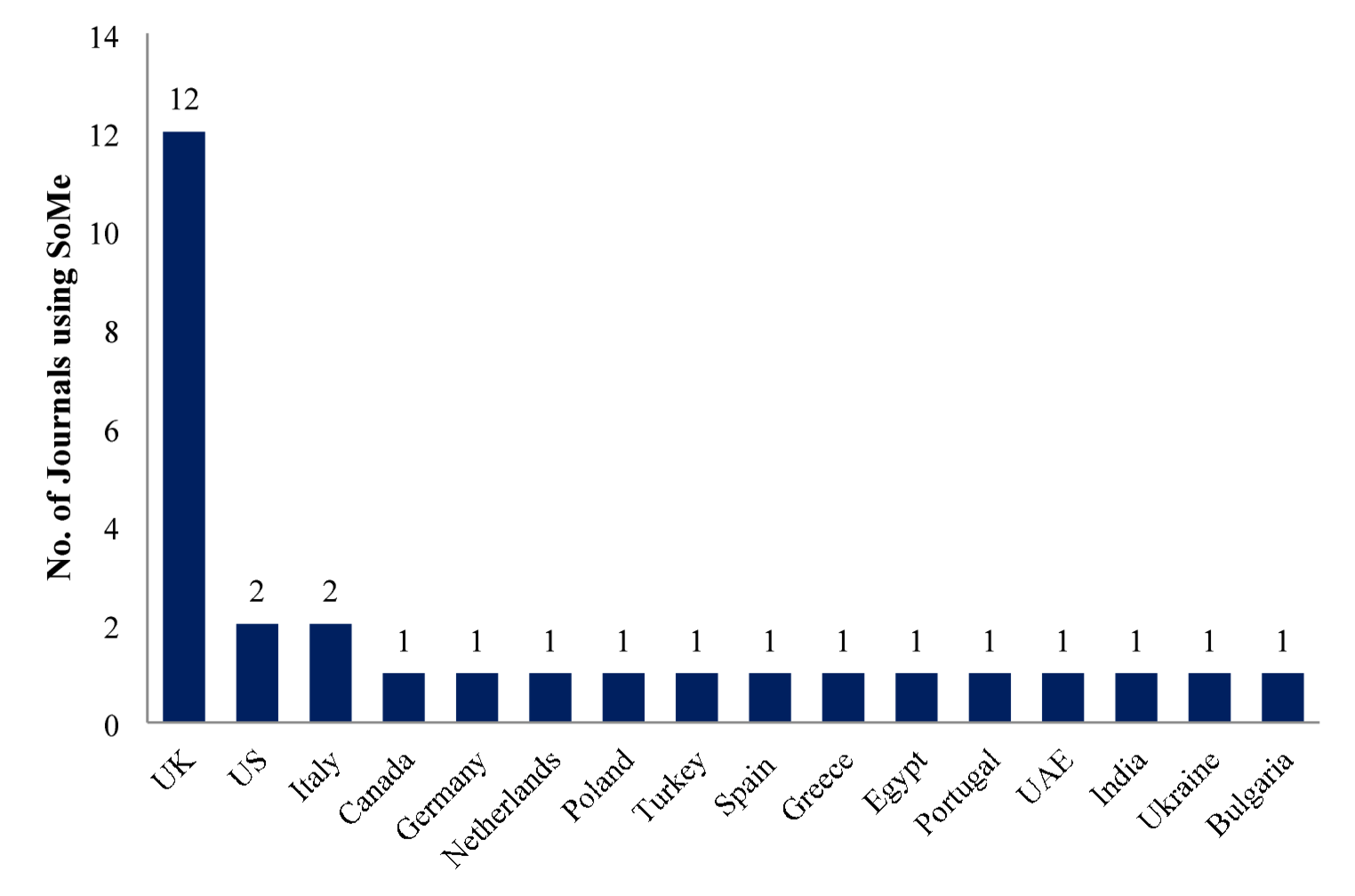
Fungal Names number: FN 822325; Fig. 1
Fig. 1
Botryosphaeria wangensis (HKAS 131605, new habitat record) a Appearance of conidiomata on the host. b, c Sections of conidiomata. d Ostiole. e Conidiomatal wall. f, g Conidiogenous cells with developing conidia. h Conidiogenous cell with conidia. i, j Conidia. Scale bars: b, c = 80 μm. d, e = 30 μm. f–j = 10 μm
Saprobic on decaying wood submerged in a freshwater lake. Asexual morph: Conidiomata 150–240 μm high, 140–200 μm wide, pycnidial, solitary or aggregated, globose to subglobose, irregular, semi-immersed to superficial, papillate, ostiolate, uniloculate, dark brown to black. Conidiomatal wall 20–45 μm thick, composed of several layers of hyaline to black brown cells of textura angularis. Conidiophores reduced to conidiogenous cells. Conidiogenous cells (4–)6–15(–19) × 2–3.6 μm (\(\bar}\)= 11 × 2.7 μm, n = 30), monophialidic, hyaline, smooth-walled, ampulliform to subcylindrical or lageniform. Conidia (15–)17–22(–25) × 4.5–6 μm (\(\bar}\)= 20 × 5.3 μm, n = 50), acrogenous, hyaline, fusiform, or irregularly fusiform, aseptate, smooth-walled, guttulate. Sexual morph: Undetermined.
Material examined: CHINA, Yunnan Province, Lijiang City, Chenghia Lake, 26° 28′ 37.16″ N, 100° 41′ 13.37″ E, on decaying submerged wood, 27 May 2021, S. Luan, H419 (HKAS 131605).
Notes: Phylogenetic analysis showed that our new collection clustered with Botryosphaeria wangensis (CERC 2298, ex-type) (Fig. 2). Morphologically, our new collection closely resembles B. wangensis with semi-immersed to superficial, unilocular conidiomata with a central ostiole; monophialidic, ampulliform to subcylindrical or lageniform, hyaline conidiogenous cells; and fusiform, or irregularly fusiform, aseptate, hyaline conidia (Li et al. 2018). However, the conidia of our new collection are slightly smaller [17–22(–25) × 5–6 μm vs. (20.5–)22–26(–29) × (4.5–)5.5–6.5(–7.5) μm] than B. wangensis (CSFF2030, holotype), which could be attributed to differences in the growth environment. The conidia morphology of our new collection was observed from natural fruiting bodies, while those of the holotype were observed from fruiting bodies produced on needles of Pinus sp. on water agar (Li et al. 2018). The nucleotide comparison between our collection, B. wangensis (HKAS 131605) and B. wangensis (CSFF2030) shows a difference of 1 bp (including 1 gap) in the ITS region. Based on morphological characteristics and phylogenetic analysis, we therefore identified the new collection as B. wangensis. The type of B. wangensis was collected from twigs of Cedrus deodara tree in Henan, China (Li et al. 2018). However, our new collection is saprophytic, isolated from a plateau freshwater lake in Yunnan, marking the first report of this species from freshwater habitats. This report thereby expands both its known distribution and ecological range.
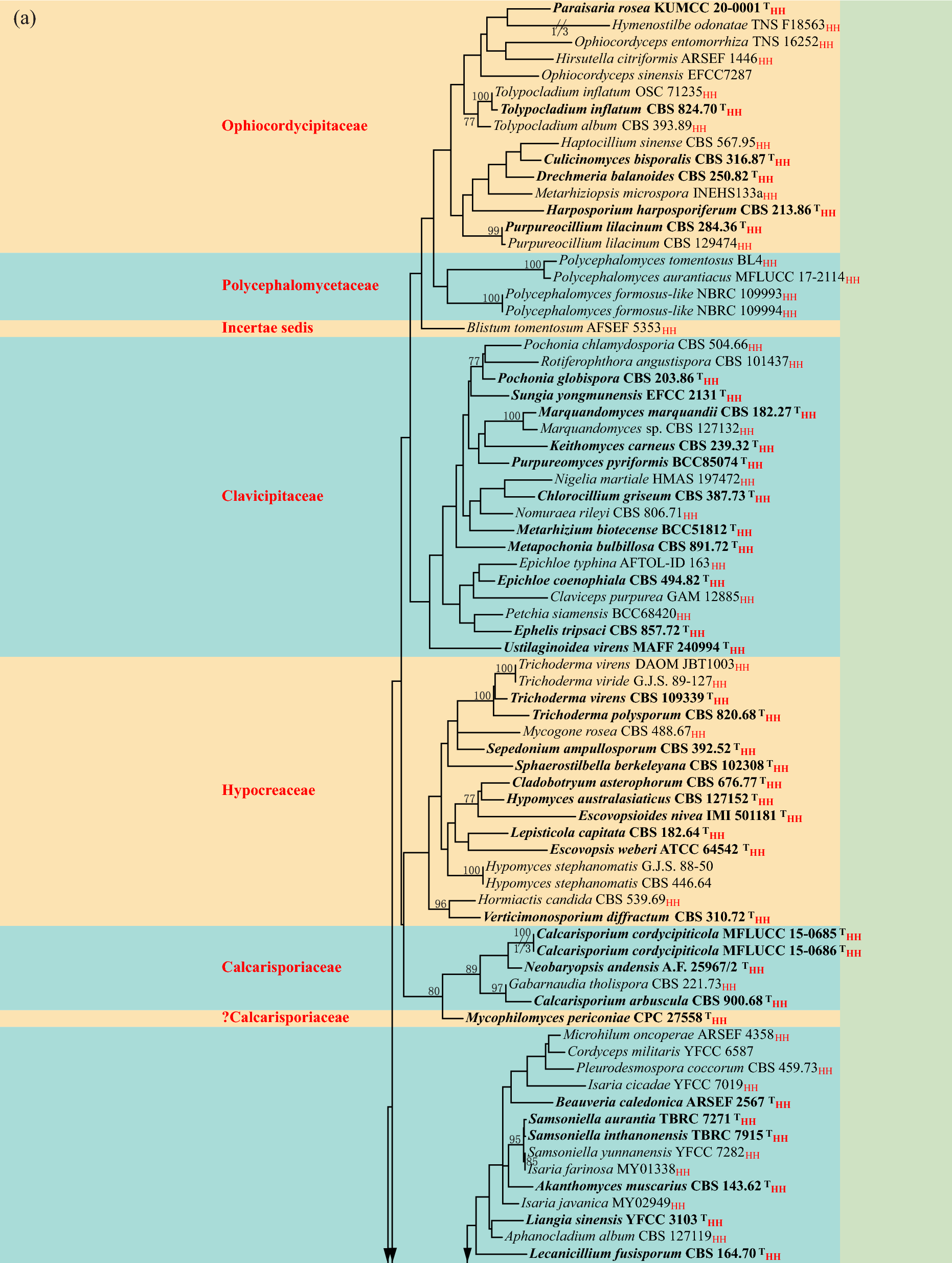
Fig. 2
Phylogram generated from maximum likelihood analysis based on combined ITS, tef1-α and tub2 sequence dataset representing Botryosphaeriaceae. Sixty-eight strains are included in the combined analyses which comprise 4682 characters including gaps (495 characters for ITS, 305 characters for tef1-α, 433 characters for tub2). Pseudofusicoccum ardesiacum (CBS 122062) and P. olivaceum (CBS 124939) were selected as the outgroup taxa. Phylogenetic trees generated from maximum likelihood and Bayesian inference analyses were similar in overall topologies. The best scoring RAxML tree with a final likelihood value of -5480.307565 is presented. The matrix had 471 distinct alignment patterns, with 9.78% undetermined characters or gaps. Estimated base frequencies were as follows: A = 0.203366, C = 0.314145, G = 0.260272, T = 0.222217; substitution rates AC = C: 0.618551, AG = 4.525254, AT = 1.594256, CG = 0.956459, CT = 7.341859, GT = 1.000000; Tree-Length = 0.759198; gamma distribution shape parameter α = 0.212336. Bayesian posterior probabilities (BYPP) from MCMC were evaluated with a final average standard deviation of split frequencies less than 0.01. Bootstrap support values for maximum likelihood (ML) greater than 60% and Bayesian posterior probabilities (BYPP) greater than 0.95 are defined above the nodes as ML/BYPP. The type strains indicated are in bold and newly generated sequence are shown in blue
Neofusicoccum Crous, Slippers & A.J.L. Phillips
Notes: Neofusicoccum was proposed by Crous et al. (2006) to accommodate species with botryosphaeria-like teleomorph and dichomera-like synanamorph, and with N. parvum (Pennycook & Samuels) Crous, Slippers & A.J.L. Phillips as the type species. Many species of this genus were transferred from Botryosphaeria, Dothiorella and Fusicoccum (Crous et al. 2006). Neofusicoccum species are commonly reported as phytopathogens and potential pathogens, associated with shoot blight, canker, and dieback of plants (Li et al. 2018, 2020a; Aiello et al. 2020; Hattori et al. 2021a; Marais et al. 2021; Zhao et al. 2022; Si et al. 2023). Some species are also saprophytes isolated from dead plant tissue (Zhang et al. 2017b; Crous et al. 2021b). A large number of species have been described in Neofusicoccum based on morphological data and phylogenetic analysis. Currently, 72 epithets recorded in the Index Fungorum database (accessed March 30, 2025). These species are described from terrestrial habitats, and there are no records from freshwater habitats. This study provides the first report of Neofusicoccum from freshwater habitats.
Neofusicoccum sichuanense X.L. Xu & C.L. Yang, Frontiers in Microbiology 13(no. 1016548): 13 (2022)
Fungal Names number: FN 845074; Fig. 3
Fig. 3
Neofusicoccum sichuanense (HKAS 131632, new habitat record) a, b Appearance of conidiomata on the host. c, d Sections of conidiomata. e–g Conidiomatal wall. h–k Conidiogenous cells with developing conidia. l–o Conidia. Scale bars: c, d = 70 μm, e–g = 30 μm, h–o = 10 μm
Saprobic on decaying wood submerged in a freshwater lake. Asexual morph: Conidiomata 140–290 μm high, 110–250 μm wide, pycnidial, solitary or aggregated, globose to subglobose, irregular, semi-immersed to superficial, erumpent, ostiolate, uniloculate, dark brown to black. Conidiomatal wall 20–70 μm thick, composed of several layers of hyaline to black brown cells of textura angularis. Conidiophores reduced to conidiogenous cells. Conidiogenous cells (10–)11–22(–27) × 3–5(–7) μm (\(\bar}\)= 16.5 × 4.3 μm, n = 30), monophialidic, hyaline, smooth-walled, ampulliform to subcylindrical or lageniform. Conidia (13–)16–19 × (5–)6–7(–8) μm (\(\bar}\)= 17 × 6.4 μm, n = 50), acrogenous, hyaline, fusiform, or irregularly fusiform, apex sub-obtuse, base truncate, aseptate, smooth-walled, guttulate. Sexual morph: detailed description, see Xu et al. (2022b).
Material examined: CHINA, Yunnan Province, Dali City, Cibihu Lake, 26° 09′ 29′′ N, 99° 56′ 47′′ E, on decaying submerged wood, 21 July 2021, L.L. Li, L1284 (HKAS 131632).
Notes: Phylogenetic analysis showed that our new collection clustered with Neofusicoccum sichuanense (SICAUCC 22-0099, ex-type) with 98% ML support (Fig. 2). Morphologically, our new collection resembles N. sichuanense in having pycnidial, globose to subglobose, irregular, semi-immersed to superficial, erumpent, unilocular conidiomata with a central ostiole; subcylindrical, hyaline conidiophores; monophialidic, hyaline, ampulliform to subcylindrical or lageniform conidiogenous cells; and hyaline, fusiform, or irregularly fusiform, aseptate conidia with sub-obtuse apex and truncate apex, and the conidial size was not different (Xu et al. 2022b). The nucleotide comparison between our collection, N. sichuanense (HKAS 131632) and N. sichuanense (SICAUCC 22-0099) showed no difference in the ITS, tef1-α and tub2 sequence. Therefore, based on the morphological characteristics and phylogenetic analysis, we identified the new collection as N. sichuanense. The holotype of N. sichuanense parasitized on the living twigs of Juglans regia in Guangyuan City, Sichuan Province. In contrast, our new collection from decaying wood submerged in a plateau lake in Yunnan Province represents the first report of this species in a freshwater habitat and expanding it’s known habitat and distribution range.
Kirschsteiniotheliales Hern.-Restr., R.F. Castañeda, Gené & Crous
Kirschsteiniotheliaceae Boonmee & K.D. Hyde
Notes: Kirschsteiniotheliaceae was introduced by Boonmee et al. (2012) to accommodate taxa grouping with Kirschsteiniothelia aethiops (Sacc.) D. Hawksw. (type species). Based on morphological data and phylogenetic analysis, K. elaterascus and K. maritima were transferred to Morosphaeriaceae and Mytilinidiaceae, respectively (Boonmee et al. 2012). Kirschsteiniotheliaceae currently contains a single genus, Kirschsteiniothelia, with more than 40 species have been reported from terrestrial and freshwater habitats (Hyde et al. 2023; Raymundo et al. 2023; Senanayake et al. 2023; Zhang et al. 2023d; de Farias et al. 2024).
Kirschsteiniothelia D. Hawksw.
Notes: Kirschsteiniothelia is the single genus in Kirschsteiniotheliaceae, introduced by Hawksworth (1985) to accommodate six species close to Microfhelia incrusfans, with K. aethiops (Berk. & Curtis) D. Hawksw. as the type. Kirschsteiniothelia was initially placed in Pleosporaceae, but subsequent multigene phylogenetic studies showed that K. aethiops was not closely related to Pleosporaceae (Hawksworth 1985; Schoch et al. 2006). Based on morphological data and phylogenetic analysis, Boonmee et al. (2012) introduced Kirschsteiniotheliaceae to accommodate species grouping with K. aethiops (Sacc.) D. Hawksw., as well as transferred K. elaterascus and K. maritime to Morosphaeriaceae and Mytilinidiaceae, respectively. The sexual morph of Kirschsteiniothelia is characterized by superficial, erumpent, globose to subglobose, membranaceous, scattered or loosely aggregated ascomata with or without central papilla; bitunicate, fissitunicate, cylindricalclavate asci; and ellipsoidal, slightly curved, septate ascospores with or without a distinct gelatinous sheath (Boonmee et al. 2012; Mehrabi et al. 2017; Hyde et al. 2018). The asexual morphs of Kirschsteiniothelia include dendryphiopsis-like and sporidesmium-like (Boonmee et al. 2012; Su et al. 2016; Bao et al. 2018; Jayawardena et al.
Comments (0)