
Remember me






GT is a promising approach for treating monogenic diseases, such as MPS IVA. Meaningful progress using viral and non-viral delivery systems has enabled approaches aimed at inducing long-term expression of GALNS. In this regard, AAV-, LV-, and CRISPR/Cas9-based GT have been evaluated for the treatment of MPS IVA using in vitro, in vivo, and ex vivo approaches. Table 2 summarizes the primary outcomes of preclinical studies for MPS IVA.
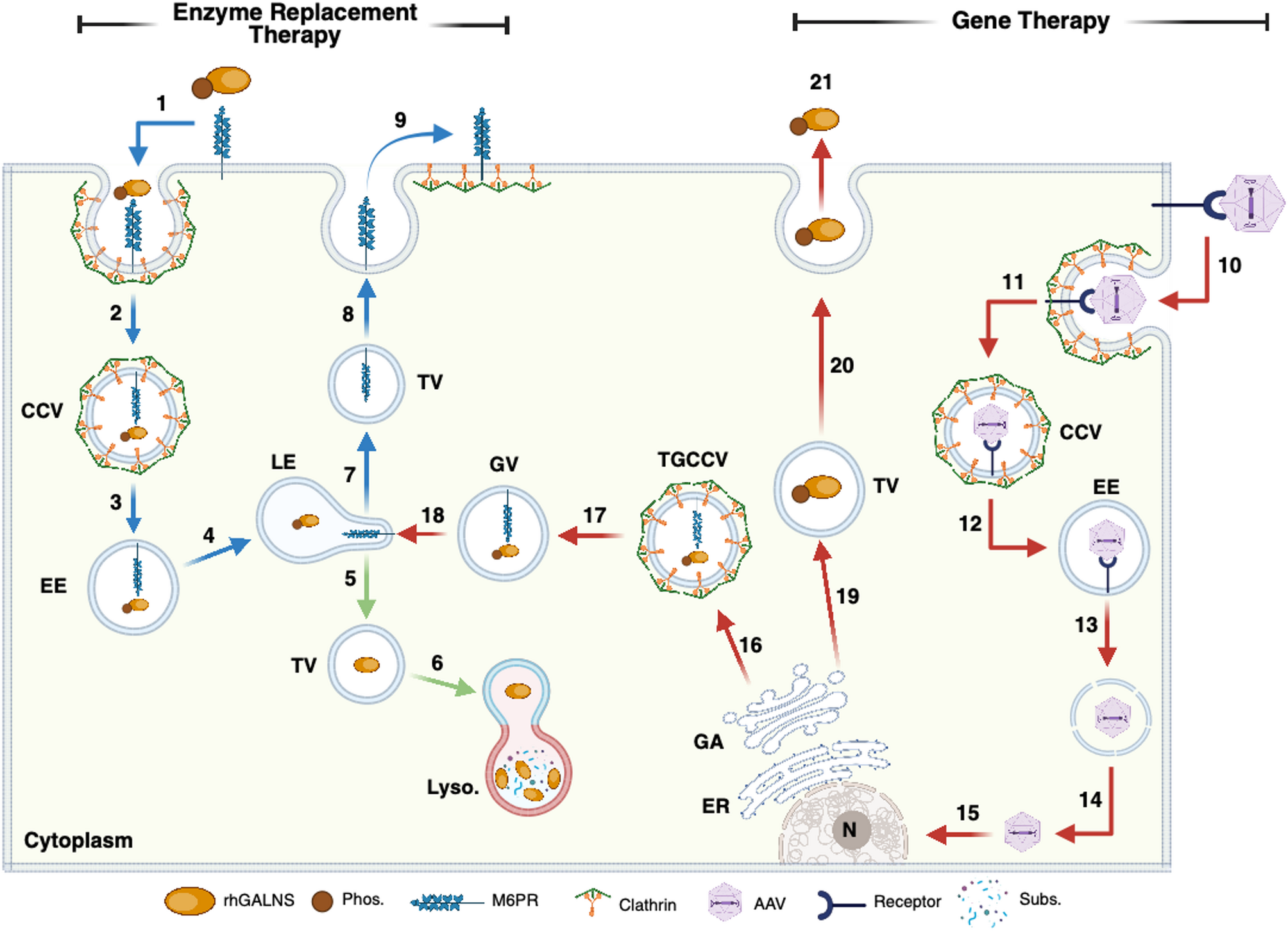
Table 2 Main findings in preclinical studies in MPS IVAAAV-based GTAdeno-associated virus (AAV)-based GT utilizes the natural ability of AAV to infect mammalian cells through receptor-mediated endocytosis [65, 66]. Upon endocytosis, viral particles reach the nucleus via endosomal escape, allowing the virus to enter the cytoplasm and subsequently the nucleus (Fig. 1). The AAV genome remains as an episome in the nucleus, where gene transcription occurs, mediated by the cell’s transcription machinery [65]. Various AAV serotypes have been identified, differing in their cell tropism, which makes them a valuable tool for GT in targeting specific tissues. Significant limitations of AAV include preexisting anti-AAV antibodies, vector dilution, limited packing capacity (4.7 kb), and potential off-target effects [67, 68]. Despite these challenges, AAV has shown promising outcomes in preclinical assessments using MPS IVA mouse models [21,22,23,24,25, 58, 59, 61,62,63, 69] and rat models [64].
Initial studies conducted by Sawamoto et al. (2020) evaluated the efficacy of AAV8 in delivering hGALNS and correcting systemic and bone manifestations in 4-week-old MPS IVA mice [58]. AAV8 was engineered to express or not the bone-targeting aspartic acid octapeptide (D8) upstream of the hGALNS cDNA sequence driven by the liver-specific promoter TGB [58]. D8 increases the vector affinity by hydroxyapatite [70]. This study demonstrated that AAV8-mediated GT results in supraphysiological levels of the hGALNS enzyme in several tissues, leading to a decrease in KS in plasma, liver, and lung. Most importantly, AAV8-GT decreases chondrocyte vacuolization in the articular cartilage and plate growth region, thus suggesting that bone pathology is ameliorated while nearly correcting heart pathology [58]. Although plasma hGALNS levels were significantly higher when mice were treated with D8-expressing hGALNS, it did not lead to a superior outcome in bone pathology recovery when compared to unmodified hGALNS [58]. Later experiments performed by Herreño-Pachón et al.. (2024) found comparable efficacy in MPS IVA mice, supporting the notion that AAV8-mediated GT is a potential strategy for treating MPS IVA [59].
The immune response raises a crucial concern in AAV-based GTs. To assess the influence of sex on the presence of anti-hGALNS neutralizing Abs and GT efficacy, Piechnik et al.. (2022) treated young male and female MPS IVA mice with AAV8 vectors carrying TGB-GALNS cDNA under the control of TGB and the ubiquitous promoter CAG [60]. In this study, the authors found that female mice elicited up to a 4.6-fold higher level of anti-AAV neutralizing Abs compared to male mice, along with a lower hGALNS enzyme activity level (6.8-fold lower) resulting from a decline in hGALNS levels over time [60]. Consistently, bone and cardiac pathology were notably improved in male mice compared to female mice [60]. Similar findings were reported regardless of the promoter driving hGALNS expression. These findings outline the importance of evaluating the sex influence in GT strategies.
A recent study published by Rintz et al.. (2023) demonstrates the potential use of AAV8 for expressing the C-type natriuretic peptide (CNP) to induce bone growth in MPS IVA mice [61]. CNP is a small bone-penetrating molecule that stimulates bone growth by interacting with the natriuretic peptide receptor B (NPR-B) in proliferative and pre-hypertrophic chondrocytes [61]. It has been evaluated for treating certain skeletal dysplasias, such as achondroplasia [71]. In the study conducted by Rintz et al.. MPS IVA mice treated with AAV8-CNP exhibited a significant increase in body length and weight compared to untreated counterparts, with notable differences observed from week 5 post-treatment onward. Significantly, AAV8-CNP treatment effectively decreases KS levels in plasma while normalizing them in bone [61]. Likewise, chondrocytes at the growth plate were less vacuolated and more organized in a columnar structure than in untreated MPS IVA mice. Bone architecture, evaluated using micro-CT, consistently revealed increased trabecular volume, decreased trabecular thickness, and an increase in total cortical medullary area [61].
Although the findings reported by Rintz et al.. support that CNP could become a potential alternative for inducing bone growth, a study published by Ago et al.. (2025) showed that N-terminal pro-C-type natriuretic peptide (NT-proCNP) is found elevated in MPS IVA patients [72], raising new concerns about the safety of the CNP overexpression through AAV-based vectors, as CNP regulates vital processes such as vascular homeostasis and blood pressure [73, 74]. In this regard, later studies conducted by Rintz et al.. (2024) showed that, despite the significant improvement in bone pathology, high AAV8-CNP doses (4 × 103 GC/Kg) result in excessive bone growth in MPS IVA mice, leading to difficulty in movement in animals [62], further suggesting that CNP dosing is critical to avoid deleterious outcomes. By decreasing the AAV8-CNP dose (1 × 1012 GC/Kg), the authors found that the combination therapy, composed of AAV9-hGALNS and AAV8-CNP, led to a significant increase in hGALNS enzyme activity and a decrease in KS in several tissues, along with amelioration of bone pathology [62]. Co-expression of GALNS and CNP in AAV9 as a single expression cassette dramatically decreases GALNS enzyme activity detection in some tissues (i.e., plasma, bone), thereby barely improving bone pathology in MPS IVA mice [62]. Globally, this study outlines the potential of combination therapies using vectors with different tropisms to express hGALNS along with key bone modulators such as CNP.
Most recently, a study conducted by Khan et al.. (2025) compared the efficacy of AAV-based GT using several promoters, including CAG, TBG, LMTP, and LBTP, in ameliorating the pathological findings in an MPS IVA [63]. In this study, the authors found that CAG (∼1100% WT levels) and TBG (∼1500% WT levels) yielded the highest GALNS enzyme activity in plasma compared to LMTP (∼300% WT levels) and LBTP (∼800% WT levels) when using AAV8 vectors after 8 weeks post-treatment [63]. When using AAV9, GALNS enzyme activity in plasma was similar in LMTP (∼1200% WT levels) and LBTP (∼900% WT levels) to that observed with AAV8-TBG and AAV8-CAG. In the femur, only AAV8-CAG and AAV9-LMTP induced supraphysiological levels of the GALNS enzyme; nevertheless, KS was normalized to WT in all vectors and promoters tested, regardless of the achieved GALNS enzyme activity [63]. AAV9-LMTP led to higher GALNS enzyme activity in muscle and bone compared to other promoters. Consistently, a significant decrease in KS levels in the muscle was observed [63]. This study highlights the therapeutic efficacy of using different serotypes and promoters.
Although most of the current data in GT have been generated in MPS IVA mouse models, the lack of a skeletal dysplasia phenotype is a significant limitation [1]. In 2021, Bertolin et al.. developed an MPS IVA rat model by inducing a point mutation (Arg388Cys) at the rat GALNS gene, which corresponds to the Arg386Cys mutation in humans [64]. The novel MPS IVA rat model recapitulates the skeletal and non-skeletal alterations expected in humans, thus providing a more accurate MPS IVA animal model. Most importantly, AAV9-based GT in MPS IVA rats resulted in vector transduction in bone, cartilage, and peripheral tissues. Consistently, the authors reported that AAV9-GALNS administration corrected bone, cardiac, and tracheal alterations in 4-week-old MPS IVA-treated rats [64]. Although the findings in this study showed the potential of AAV9 in reaching and transducing avascular regions in the bone, the authors used a rat GALNS cDNA, which lacked evidence of these promising findings when using hGALNS cDNA. Rat-derived anti-hGALNS neutralizing Abs could decrease the therapeutic response as observed in mouse models [63]. Humanized rats (i.e., expressing truncated hGALNS) could offer a novel approach to testing the therapeutic efficacy of AAV9-hGALNS in this innovative MPS IVA animal model.
LV-based GTLentiviruses, a subclass of retroviruses, have gained particular interest due to their ability to integrate into the host genome and transduce both dividing and non-dividing cells [75]. Originally derived from the human immunodeficiency virus (HIV-1), lentiviral vectors (LVs) have been extensively engineered for enhanced biosafety and transduction efficiency, leading to their adoption in various clinical trials, including those for primary immunodeficiencies and hemoglobinopathies [76, 77]. LV-mediated transgene delivery offers key advantages over AAV, as it has a larger packaging capacity and a lower immunogenic profile [75, 77]. Nevertheless, insertional mutagenesis, random integration, and epigenetic silencing remain the most significant challenges [78]. In MPS IVA, several in vitro and ex vivo approaches have evaluated the efficacy of LV-mediated GT.
Initial studies performed by Puentes-Tellez et al.. (2021) demonstrated the suitability of HIV-derived LVs for transducing MPS IVA fibroblasts [79]. Upon transduction with LVs carrying hGALNS cDNA alone or in combination with hSUMF1 cDNA, a significant increase in the GALNS enzyme activity, a decrease in the lysosomal mass, and autophagy flux recovery were noticed [79]. Mutation-causing GALNS deficiency influenced the successful recovery of biomarkers in MPS IVA fibroblasts [79], suggesting that some mutations may lead to differential responses in LV-based GT approaches.
Most recent studies by Celik et al.. (2024) evaluated, for the first time, the use of LV in an ex vivo approach using mouse HSCs [21]. In this study, the authors aimed to test the efficacy of LVs carrying hGALNS expression cassettes under the control of ubiquitous (CBh) or cartilage-specific (COL2A1) promoters. LVs were used to transduce murine HSCs, which were then transplanted into busulfan-treated MPS IVA mice [21]. Interestingly, both vectors led to supraphysiological hGALNS activity in white blood cells (WBC) from treated MPS IVA mice, suggesting that LV-mediated HSC transduction does not affect the in vivo differentiation of HSC into mature blood circulating cells [21]. Besides, LV-CBh-hGALNS-treated mice showed detectable levels of the GALNS enzyme in the tibia, while LV-COL2A1-hGALNS-treated mice did not. Interestingly, in LV-COL2A1-hGALNS-treated mice the KS levels were normalized to WT levels, while LV-CBh-hGALNS-treated mice showed a non-significant KS decreased in bone (humerus) [21]. Similarly, LV-COL2A1-hGALNS treatment completely rescued the heart pathology, whereas LV-CBh-hGALNS failed to recover the cell vacuolization in the heart to the same extent as LV-COL2A1-hGALNS, although a slight improvement was observed [21]. LV-COL2A1-hGALNS and LV-CBh-hGALNS partially corrected bone pathology. Additionally, a non-significant improvement in bone microarchitecture was observed in MPS IVA mice treated with COL2A1-hGALNS [21]. Although ex vivo LV-based GT still needs to be optimized to achieve complete bone correction, including bone microarchitecture, this pioneering study offers proof-of-concept for investigating ex vivo GT in the treatment of MPS IVA.
CRISPR/Cas9-based GTThe CRISPR/Cas9 system enables gene modification, including gene knockout, knocking and base editing [80, 81]. Unlike traditional GT using AAV and LV, the CRISPR/Cas9 system enables the direct correction of pathogenic mutations at their native loci, potentially restoring normal gene function with long-term durability [80]. Most promising, the CRISPR/Cas9 system allows the insertion of long-expression cassettes at safe harbors [82, 83], resulting in the recovery of gene expression regardless of the disease-causing mutation. This is particularly interesting for MPS IVA, as more than 350 mutations have been identified in the GALNS gene [1]. The CRISPR/Cas9 system, primarily utilizing Cas9 nickase (nCas9), has been tested in MPS IVA fibroblasts [24, 25, 69] and CD34 + cells [22], as well as in MPS IVA mouse models [23], using both non-viral vectors (NVVs) and viral vectors (VVs). Figure 2 shows a comparison between wtCas9 and nCas9, which have been evaluated in models of MPS IVA.
Fig. 2
CRISPR/Cas9 alternatives evaluated MPS IVA models. Key features of Cas9 wild-type (wtCas9) and nickase (nCas9), including preferred DNA repair pathways, base/prime editing compatibility, off-target potential, design complexity, and relevance for multi-gene editing. Applications in MPS IVA are highlighted, showing the broader utility of nCas9 in both fibroblasts and hematopoietic cells, HDR Homologous repair, NHEJ Non-homologous end-joining. sgRNA. Single guide RNA. This figure was created with Biorender.com
In vitro, Leal and Alméciga (2022) demonstrated, for the first time, the efficacy of a CRISPR/Cas9 system to insert an expression cassette carrying hGALNS cDNA at the AAVS1 locus [24]. The AAVS1 locus is a safe insertion site that enables the knock-in of foreign DNA without causing cell disturbances [84]. In the studies conducted by Leal & Alméciga (2022), the authors employed a Cas9 variant termed Cas9 nickase (nCas9), as it has been shown to decrease the potential for off-targeting while preserving its ability to induce high knock-in efficiencies [85, 86]. Consistently, DSB-mediated nCas9 was as high as 37%, while unwanted Cas9 cutting was not observed in 10 in silico-predicted off-target sequences [24]. Interestingly, 30 days post-treatment, the authors found a recovery of up to 40% in GALNS enzyme activity, which was sufficient to normalize lysosomal mass, mitochondrial-dependent oxidative stress, and global GAG accumulation [24], providing novel evidence of the CRISPR/nCas9 efficacy in treating MPS IVA. Recent studies published by Suarez et al.. (2025) further support the suitability of the CRISPR/Cas9 system for recovering the MPS IVA fibroblast phenotype [69].
A later study, conducted by Leal et al.. (2022), aimed to evaluate the transport and delivery of the CRISPR/Cas9 system using iron oxide-based nanoparticles (IONPs) [25]. In this study, the authors found that IONPs yield comparable phenotype recovery in human MPS IVA fibroblasts carrying different GALNS-affecting mutations, compared to the phenotype recovery observed when using lipofectamine [25], supporting the notion that NVVs could be a novel alternative to VVs for delivering the CRISPR/Cas9 system.
Moving towards a preclinical assessment, the CRISPR/nCas9 system was also administered to MPS IVA mice using IONPs [23]. Initial experiments demonstrated that IONPs accumulated primarily in the liver, followed by the lungs, arms, legs, spleen, and kidneys, suggesting a widespread biodistribution [23]. Liver, renal, and iron homeostasis showed no toxicity related to IONP administration, indicating the dose used (2.5 µg IONPs carrying 3.8 µg CRISPR/Cas9 plasmids) was safe for animals [23]. The CRISPR/Cas9 system led to a 28% recovery of GALNS enzyme activity in the plasma of treated MPS IVA mice. Significant GALNS enzyme activity increase was also reported in the liver, muscle, spleen, heart, kidneys, and lungs. Conversely, a slight, non-significant increase in GALNS was observed in the tibia and trachea [23]. Biochemical correction was observed in plasma, liver, and humerus, suggesting that non-supraphysiological GALNS levels can mediate phenotype correction as observed in classical AAV- and LV-mediated GT [58]. Despite the significant decrease in KS in the bone, the vacuolization of the chondrocytes was only partially corrected [23], further highlighting the need for strategies targeting the avascular zones of the bone that remain unaddressed.
Although in vivo assessment of the CRISPR/Cas9 has shown encouraging results, the preexisting immune response against Cas9 proteins [87] could limit its therapeutic efficacy. A study conducted by Herreño-Pachón et al. (2025) aimed to evaluate the GE efficacy of the CRISPR/nCas9 system for modifying HSCs [22]. CRISPR/nCas9-based knock-in did not affect HSCs’ properties and led to supraphysiological GALNS expression [22]. The authors also observed an increase in intracellular GALNS in MPS IVA cells upon co-culture with edited HSCs. Normalization of classical MPS IVA biomarkers was observed in MPS IVA cells [22], suggesting cross-correction. The transplantation of CRISPR/nCas9-edited CD34 + cells into MPS IVA mouse models still needs to be conducted.
Comments (0)