
Remember me
The α-mangostin has a wide range of medicinal properties, including antioxidant [23], antineoplastic [30], anti-inflammatory [31, 32], antihistamine [33], antibacterial [34], antifungal [35], antiviral [36,37,38,39,40,41,42,43,44], antimalarial [45], antidiabetic [46], antihyperlipidemic [46], cardioprotective [47, 48], hepatoprotective [49], neuroprotective [50], and immunomodulatory activities [51], among others.
In terms of its antineoplastic properties, α-mangostin has shown chemopreventive and antitumoral effects against various cancers, including cholangiocarcinoma [52], pheochromocytoma [53], glioblastoma [54], osteosarcoma [55], head and neck cancer [56], prostate cancer [57, 58], gastric cancer [59], pancreatic cancer [60,61,62], lung cancer [63], cervical cancer [43, 64], colon cancer [65], ovarian cancer [66, 67], skin cancer [68], renal cancer [69], and breast cancer [3, 70,71,72,73,74,75,76,77,78,79].
3.1 Mechanisms of α-mangostin involved in inhibiting breast cancer cell proliferationIn the context of breast cancer, while personalized therapeutic strategies have been developed and applied based on specific tumor phenotypes, α-mangostin has shown antiproliferative effects regardless of the tumor molecular expression profile. This includes its effectiveness in breast cancer cells representing tumor phenotypes such as Estrogen Receptor-positive (ER +), Human Epidermal Growth Factor Receptor Type 2 (HER-2) enriched, and Triple Negative (TN), which lacks expression of ER, Progesterone Receptor (PR), and HER-2 (Table 2).
Table 2 Antiproliferative effects of α-mangostin or mangosteen extract on breast cancer cell linesOn the other hand, α-mangostin has demonstrated the ability to suppress breast cancer cell growth and the development of pre-neoplastic lesions induced by 7,12-dimethylbenzo (α) anthracene in a mouse mammary organ culture assay, with an IC50 of 2.44 µM [23]. Numerous studies highlight the impact of α-mangostin on breast cancer, identifying several mechanisms through which it may exert its antiproliferative effects (Fig. 3).
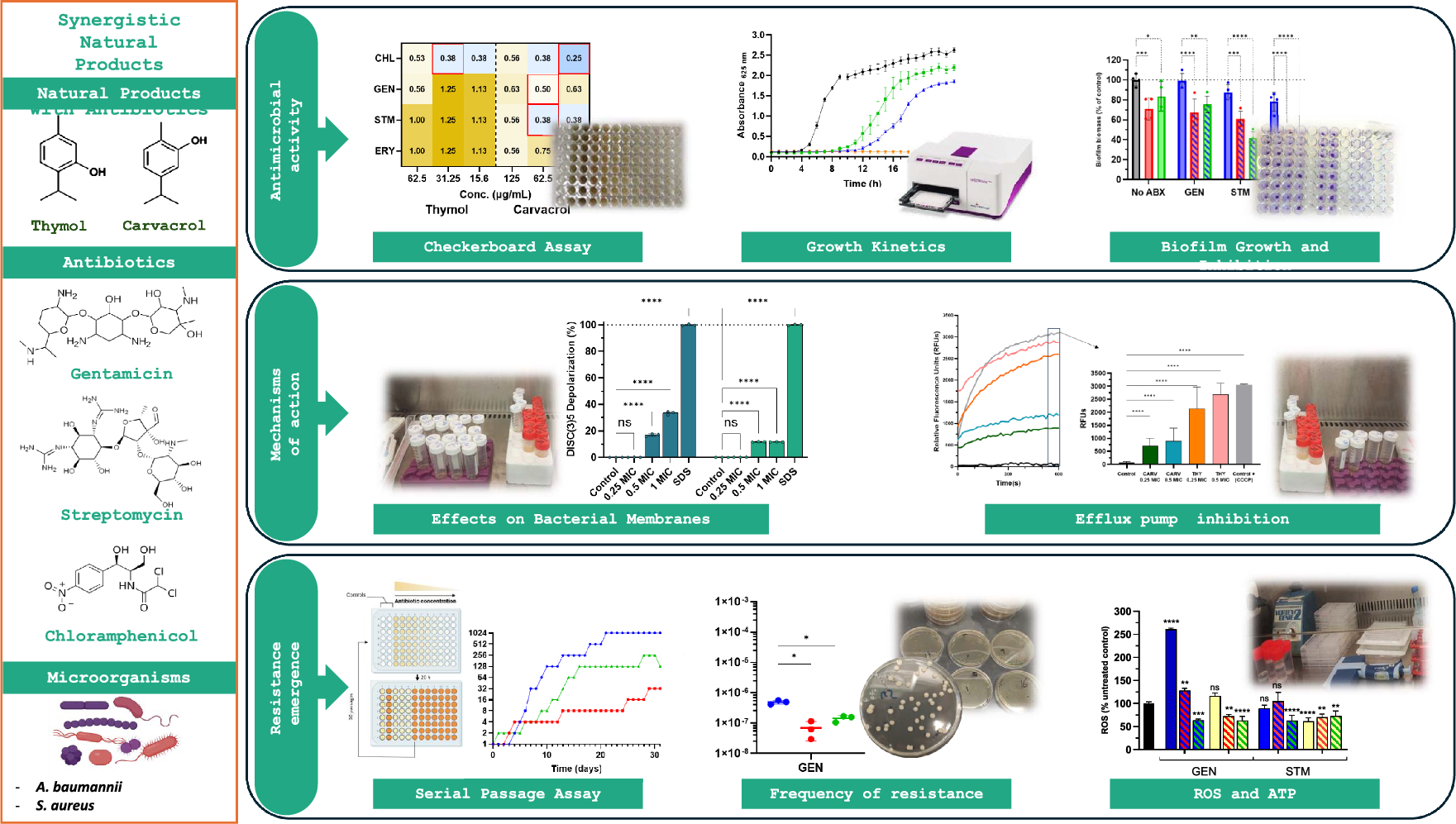
Fig. 3
Antiproliferative mechanisms of α-mangostin. α-Mangostin (AM) exhibits diverse mechanisms to inhibit breast cancer cell proliferation, including: (1) Hormonal receptor modulation: AM reduces ERα expression and phosphorylation, increases ERβ levels, and inhibits estrogen-dependent gene expression. Docking studies indicate strong interactions with ERs, and derivatives of AM act as potential ER antagonists. (2) Blockade of growth factor receptor signaling: AM inhibits HER-2 phosphorylation and EGFR, and VEGFR binding, downregulating key oncogenic pathways (RAS/RAF1/MEK/ERK and PI3K/AKT). (3) Histamine H1 receptor (H1R) inhibition: AM blocks H1R, linked to tumor growth, and reduces breast cancer histamine levels. (4) Enzyme inhibition: AM targets aromatase, topoisomerases, DNA polymerases, Lysine-Specific Demethylase 1 (LSD1), and fatty acid synthase (FAS), disrupting proliferation, DNA processes, and lipid synthesis. (5) Ion channel modulation: AM inhibits oncogenic potassium channels (KCNH1, Kv1.3, KCa3.1), calcium transport (ORAI1, Ca2+ ATPase), impacting cell signaling and viability. (6) Cell cycle modulation: AM induces G1-phase arrest and downregulates cyclin-CDK complexes, preventing uncontrolled cell division. Figure created using BioRender.com
3.1.1 Modulation of hormone receptors by α-mangostin in breast cancerIn the canonical mechanism of action, ERα functions as a transcription factor upon ligand binding, driving the transcription of genes involved in cell proliferation. One mechanism through which α-mangostin inhibits cancer cell proliferation is by inhibiting ERα (Fig. 3). Specifically, at a concentration of 10 µM, α-mangostin suppresses ERα protein expression and decreases the estrogen-dependent gene Ps2 in MCF-7 cells. Notably, siRNA-mediated knockdown of ERα levels, followed by subsequent treatment with α-mangostin, reduces its effects, suggesting that the xanthone exerts its activity, at least in part, through ERα [76]. Additionally, ERα phosphorylation is reduced in T-47D cells treated with 30 µM of α-mangostin, suggesting that α-mangostin may additionally inhibit cell proliferation and survival by suppressing ERα signaling pathway [74]. Moreover, treatment of MCF-7 cells with mangosteen rind extract (50 and 100 µg/mL) decreased ERα gene expression in a concentration-dependent manner while increasing ERβ levels [80]. ERβ, in particular, plays a significant role in breast cancer, often acting as a tumor suppressor; while ERα promotes cancer cell proliferation, ERβ tends to inhibit it, and high levels of this protein are associated with better prognosis and improved survival rates [90].
Given the significance of estrogen signaling in breast cancer progression, blocking this pathway is a key therapeutic strategy in ER-positive tumors. In this context, it has been demonstrated that α-mangostin at 3.5 µM reduces the expression of estrogen-regulated genes, such as those encoding prolactin and PR, in MCF-7 cells, suggesting the interference with estrogen signaling [4].
In an additional study, the anti-proliferative effects of four xanthones, including α-mangostin, β-mangostin, mangostenol, mangaxanthone B, along with three benzophenones and one sterol, were evaluated. Initially, the antiproliferative activity of these compounds was assessed in the breast cancer cell lines MCF-7 and MDA-MB-231, identifying α-mangostin as the most potent compound, with IC50 values of 4.43 µM and 3.59 µM, respectively. Subsequently, a molecular docking simulation study was conducted to evaluate the interactions of xanthones with 2OCF (Protein Data Bank ID for the protein structure of ER). The analysis regarding α-mangostin revealed key interactions contributing to its potential biological activity. Specifically, the prenyl group at C8 of α-mangostin interacts with the Leu320, Trp393, Phe445, and Val446 residues of 2OCF at various distances (Å) through hydrophobic interactions. The aromatic ring A of α-mangostin interacts with Arg394, while rings B and C with Ile326 and Arg394 through hydrophobic interactions. Furthermore, the hydroxyl group at C1 and the carbonyl group (C = O) interact with Trp393 through hydrogen bonding. Additionally, a molecular docking simulation of xanthones was performed with 4PIV (Protein Data Bank ID for the β-ketoacyl part of Fatty Acid Synthase, FAS), which will be discussed in detail in the section regarding FAS inhibition [19].
Other studies have also explored α-mangostin and its derivatives as potential ERα antagonists. In one of them, α-mangostin was chemically modified by substituting the hydroxyl group at C6 with benzoyl derivatives. Computational analyses predicted that the resulting derivatives AMB-1, AMB-2, and AMB-10 may function as potential ERα antagonists. Of note, these findings were based solely in silico approaches and were not validated through experimental assays [71]. More recently, another study explored the conjugation of glycine to the hydroxyl groups at positions C3 and/or C6 of the α-mangostin, leading to the development of glycine-conjugated α-mangostin derivatives. Among these, the Am1Gly, with glycine attached to the hydroxyl group at C3, demonstrated promising potential as an ERα antagonist. Further pharmacophore modeling revealed that Am1Gly1 and Am2Gly2 possess structural features comparable to those of active drugs targeting ERα, reinforcing their potential for therapeutic development. Molecular docking analyses indicated that Am1Gly1 interacts with key residues essential for functioning as a 4-hydroxytamoxifen-like ERα antagonist, exhibiting dynamic interactions similar to those observed with 4-hydroxytamoxifen. These findings were supported by molecular dynamics simulations, which confirmed consistent amino acid variation patters over 200 ns. Additionally, binding affinity analysis, molecular docking, dynamic simulations, and pharmacophore modeling collectively supported the potential of Am1Gly1 as an ERα antagonist. This study was not accompanied by experimental validation [91].
3.1.2 Inhibition of growth-factor receptors signaling by α-mangostin in breast cancerHER-2 is a receptor with intrinsic tyrosine kinase activity, comprising an extracellular ligand-binding domain, a transmembrane domain, and an intracellular tyrosine kinase domain. While HER-2 does not have a known ligand that directly binds to it, it acts as the preferred heterodimerization partner for other members of the epidermal growth factor receptor (EGFR) family (ErbB). Thus, upon ligand binding to other ErbB members, HER-2 is recruited, leading to the autophosphorylation of its intracellular tyrosine kinase domains. This activation triggers the phosphatidylinositol 3-kinase (PI3K)/protein kinase B (PKB, also known as AKT) and mitogen-activated protein kinase (MAPK) signaling pathways, which are crucial for processes closely related to breast tumorigenesis such as cell survival, proliferation, differentiation, and apoptosis [92]. Notably, treatment of T-47D breast cancer cells with 30 µM of α-mangostin resulted in reduced phosphorylation of HER-2 at the Tyr1221/1222 within 30 min, leading to the deactivation of RAS/RAF1/MEK/ERK and PI3K/AKT signaling pathways, and consequently, an anti-proliferative and pro-apoptotic effect (Fig. 3) [74].
Additionally, molecular docking analysis provided evidence that garcinone E, another prenylated xanthone, which differs from α-mangostin by an isoprenyl group at the C5 position and hydroxyl group at the C7 position (Fig. 2), binds tightly to the EGFR and the vascular endothelial growth factor receptor (VEGFR), inhibiting their kinase activity with IC50 values of 315.4 nM and 158.2 nM, respectively. Furthermore, garcinone E blocks the endothelial growth factor (EGF)- and vascular endothelial growth factor (VEGF)-induced phosphorylation of EGFR and VEGFR, respectively. Although α-mangostin also binds to these receptors, it does so with less affinity than garcinone E. Beyond its molecular interactions, garcinone E demonstrated potent anti-cancer activity, inhibiting cancer cell proliferation, endothelial cell migration, invasion, and tube formation. Its dose-dependent suppression of angiogenesis was confirmed in ex vivo and in vivo models. In an MDA-MB-231 xenograft model, garcinone E (2 mg/kg) significantly reduced tumor growth, weight, and microvessel density, correlating with downregulation of VEGFR2, EGFR, and Ki-67 [93]. However, mangosteen rind extract (50 and 100 µg/mL) decreased the gene expression of EGFR in MCF-7 cells in a concentration-dependent manner [80]. In the TN breast cancer phenotype, EGFR is highly expressed, which is associated with cancer progression, proliferation, metastasis, and drug resistance, making EGFR a promising pharmacological target [94]. Therefore, α-mangostin could be considered a potential compound for targeting this receptor.
3.1.3 Inhibition of H1R by α-mangostin in breast cancerα-Mangostin has been identified as an inhibitor of the H1R. Specifically, it exhibits a concentration-dependent inhibition of [3H]mepyramine binding, a selective H1 receptor antagonist, in rat aortic smooth muscle cells. Kinetic analysis indicates that α-mangostin acts as a competitive inhibitor of the H1R, preventing [3H]mepyramine from binding effectively with an IC50 value of 2.27 µM. Scatchard plot analysis further revealed that [3H]mepyramine binds to a high affinity receptor site with a dissociation constant (Kd) of 11.72 nM and a maximum binding capacity (Bmax) of 275.95 fmol/mg. Treatment with α-mangostin increased the Kd value to 38.02 nM, indicating reduced affinity, while the Bmax value remained unchanged. These findings confirm that α-mangostin function as a specific H1R antagonist [33] (Fig. 3). This is relevant because there is a direct correlation between endogenous histamine levels and breast cancer development. In particular, higher histamine concentrations have been observed in tumors and adjacent tissue of breast cancer patients compared to the unaffected tissue of healthy individuals [95]. Additionally, there is a significant increase in histamine levels in the plasma and tissues of patients with ductal breast cancer. The plasma histamine concentrations in women with ductal breast carcinoma depend on the number of involved lymph nodes and the grade of histologic malignancy [96]. Additionally, H1R is overexpressed in basal-type and HER-2-enriched breast tumors, which correlates with a shorter overall survival [97]. This supports the idea that histamine and its receptors are involved in cell growth. Furthermore, several antihistamines have been shown to inhibit breast cancer cell proliferation both in vitro and in vivo [98,99,100,101].
3.1.4 Inhibition of enzymatic activities associated with breast cancer cell proliferation by α-mangostin3.1.4.1 Aromatase.Aromatase plays a pivotal role in converting androgens into estrogen through aromatization. Notably, breast cancer tissues exhibit higher levels of aromatase compared to healthy tissues [102]. Consequently, one of the therapeutic strategies in the clinical practice involves aromatase inhibitors to reduce breast cancer growth. These inhibitors hamper the enzyme’s function, affecting estrogen synthesis and, consequently, blocking breast cancer cell proliferation. Interestingly, a non-cellular, enzyme-based microsomal assay, revealed that garcinone D, garcinone E, α-mangostin, and γ-mangostin exhibited a dose-dependent aromatase inhibitory activity. Among these compounds, garcinone D (IC50 = 5.2 µM) and γ-mangostin (IC50 = 6.9 µM) demonstrated the strongest inhibitory effects in this assay. In contrast, the two other xanthones, α-mangostin (IC50 = 20.7 µM) and garcinone E (IC50 = 25.1 µM), displayed moderate inhibition of aromatase activity [103] (Fig. 3).
3.1.4.2 DNA polymerases and topoisomerases.DNA polymerase synthesizes new DNA strands during replication, while topoisomerases I and II play critical roles in cutting single-stranded or double-stranded DNA to relieve tension during replication or transcription events [104]. Interestingly, α-mangostin exhibits a strong inhibitory effect on both DNA polymerases (IC50 values 14.8 µM–25.6 µM) and topoisomerases I and II (15.0 µM and 7.5 µM, respectively). Notably, α-mangostin completely suppresses the nicking activity of topoisomerase I at concentrations greater than 20 µM, while at 10 µM, it fully inhibits topoisomerase II. Interestingly, as determined by thermal transition analysis, α-mangostin does not directly bind to double-stranded DNA. This suggests that the decreased proliferation observed with α-mangostin may be partially due to the inhibition of these enzymes [105] (Fig. 3).
3.1.4.3 Lysine-specific demethylase 1 (LSD1).LSD1 is an enzyme that plays a crucial role in gene expression regulation through histone modification. It is frequently overexpressed in several human cancers, including breast cancer, where it promotes tumor progression by supporting cancer cell survival and creating a pro-oncogenic environment. As a histone demethylase, LSD1 removes methyl groups from specific lysine residues on histone proteins, mainly histone H3 at lysine 4 (H3K4) and lysine 9 (H3K9). This chromatin remodeling process, directly influences transcriptional activity, either repressing or activating gene expression [106]. In ERα-positive breast cancer, LSD1 demethylates H3K9 at ERα target genes in an estrogen-dependent manner, facilitating transcriptional activation. This regulatory mechanism is reinforced by Proline, Glutamate, and Leucine Rich Protein 1 (PELP1), a co-activator that links LSD1 to ERα, forming the LSD1-PELP1-ERα axis implicated in hormone resistance. Additionally, LSD1 indirectly promotes CYP19A1 expression, encoding aromatase, through its interaction with HER-2 signaling [107]. Conversely, LSD1 demethylates non-histone proteins, such as p53, leading to the repression of its activity [
Comments (0)