
Remember me






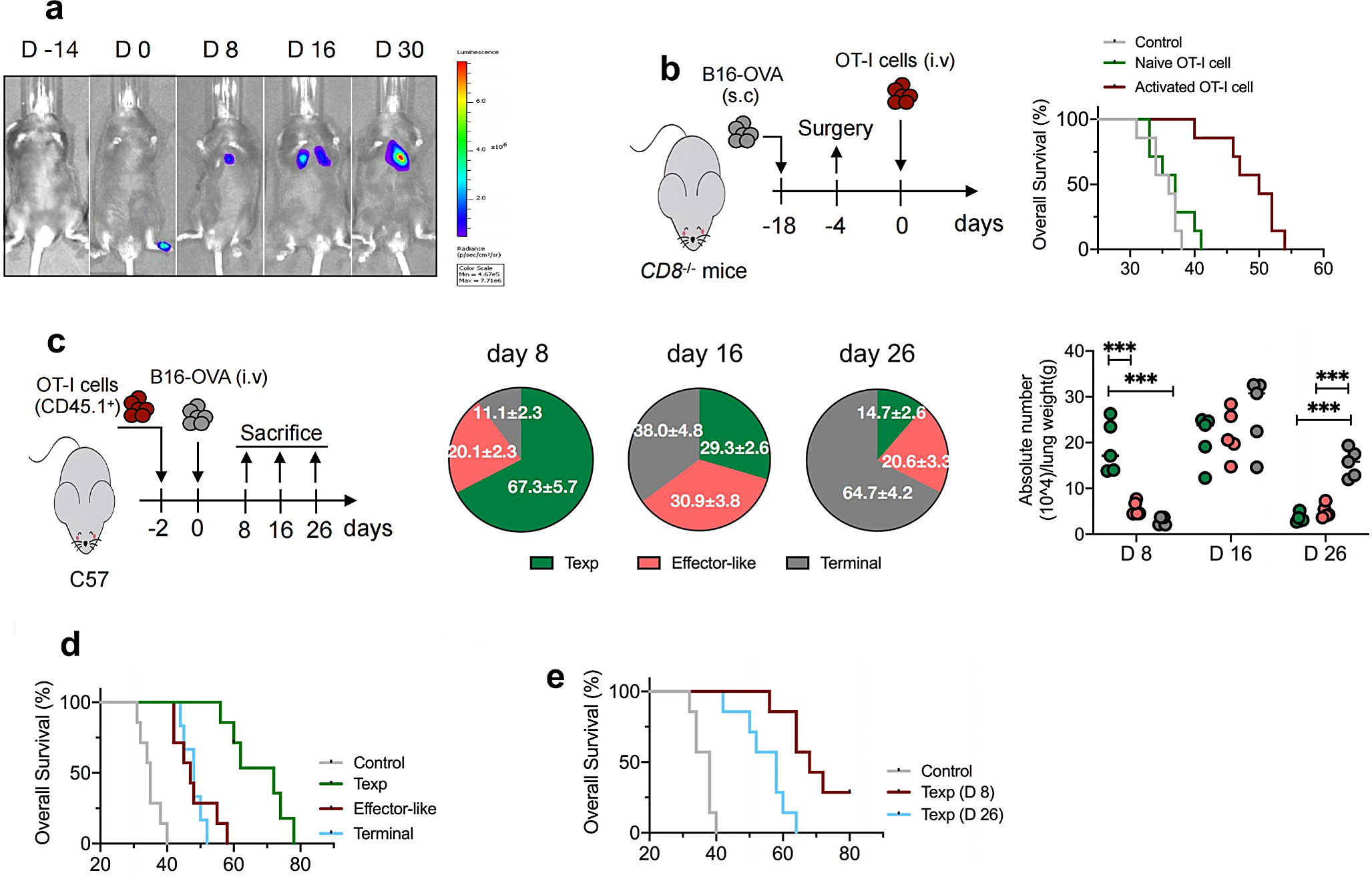
To investigate the response of tumor-infiltrating CTLs to metastatic tumors, we used an experimental mouse model of lung melanoma metastasis [18]. B16luc cells (harboring a luciferase-coding gene) were inoculated into footpad of mice and subsequent metastatic foci in the lungs were observed in proportion to the intensity of the luciferase reporter signal (Fig. 1a). Consequently, we aimed to investigate whether the adoptive transfer of antigen-specific CD8+ T cells could mitigate metastatic tumors. Prior to surgical resection of the primary tumor, mice with confirmed lung metastases were subjected to a luciferase assay. To eliminate the potential influence of endogenous CD8+ T cells, we selected CD8+ T cell-depleted (Cd8−/−) mice as hosts. Despite this, the fluorescence signal intensity in the lung tissue of Cd8−/− mice did not significantly differ from that of wild-type mice at various time points (supplemental figure S1a). A total of 10 × 105 naïve or activated splenic OT-I cells (OVA257 − 264 I-Ab- specific) were adoptively transferred intravenously into Cd8−/− mice bearing lung metastases (Fig. 1b). Mice receiving activated OT-I cells instead of naïve cells showed significantly enhanced survival rates compared with controls, indicating that the transfer of activated antigen-specific CD8+ T cells is highly efficacious in controlling metastasis.
Fig. 1
Differentiation pattern of OT-I cells within metastatic tissues and therapeutic outcome of ACT (a). Approximately 5 × 105 of B16luc cells expressed luciferase were subcutaneously injected into the footpad of C57BL/6J mice to develop the spontaneous lung metastasis mice model, luciferin was administrated on day 0, 8, 16, 30, and light emission was recorded. Representative bioluminescence images are shown. (b). Cd8−/− mice were subcutaneously injected with B16-OVA cells, after 14 days, lung-metastasis mice were confirmed by luciferase assay prior to surgery. Totally 1 × 106 of naïve splenic OT-I cells or activated OT-I cells that were derived from stimulation with anti-CD3/CD28 and IL-2 in vitro were i.v adoptively transferred into mice. No treatment mice were used as control. Survival curve of mice was calculated. Red line versus gray line, p < 0.01; Green versus gray line, p = 0.33; log rank test was used. (c). Naïve splenic OT-I cells (5 × 105, CD45.1+) were transferred into C57BL/6J mice (CD45.2+, n = 5 per group) subsequently injection with B16-OVA cells (3 × 105) via tail vein. On day 8, 16 and 26 post transfer, lungs were harvested and tissue-infiltrating lymphocytes (TILs) were isolated and subjected to flow cytometry. Frequency and absolute number of precursor-exhausted (Texp), effector-like and terminal Tex subsets of OT-I cells were measured. Statistical comparisons were performed using an unpaired t-test, ***p < 0.001. Data are shown as the mean ± s.e.m. (d). Approximately 1 × 106 of lung-derived Tex cells that were sorted from donor mice on day 12 as described in (c), or isolated Texp cells on day 8 or day 26 (e), were adoptively transferred into pre-established metastasis-bearing Cd8−/− mice 4 days post-surgery. Survival curve of mice was calculated. In (d), Blue / red line versus gray line, p < 0.01; Green versus blue / red line, p < 0.01; In (e), Blue line versus gray line, p < 0.01; Red line versus blue line, p = 0.015. log rank test was used
The differentiation pattern of antigen-specific CD8+ T cells in the tumor microenvironment has been previously investigated [11]. However, there is a limited understanding of this phenomenon in the context of metastatic tissue. A total of 5 × 105 splenic CD45.1+OT-I cells were intravenously administered to mice on day − 2, followed by intravenous inoculation with B16-OVA cells (B16 cells expressing the OVA257 − 264 peptide). Subsequently, the lung tissue was harvested, and the infiltrated CD44+OT-I cells were analyzed using flow cytometry. Whole tissue-infiltrating OT-I cells were classified into Texp cells (TCF1+Tim3− CX3CR1−), transitory effector-like Tex cells (TCF1−CX3CR1+), and terminal Tex cells (TCF1−Tim3+CD101+) (supplemental figure S1b). Notably, we observed an initial predominance of Texp cells among expanded antigen-specific CD8+ T cells, which gradually declined thereafter. In addition, there was a 3- to 5-fold increase in the proportion of terminal Tex cells at the examined time points, whereas the effector-like Tex cell population exhibited only marginal fluctuations during metastatic progression (Fig. 1c). These findings demonstrated that tumor-specific CD8+ T cells exhibit high heterogeneity in exhaustion within metastatic tissues.
Co-expression of TCF1 and Slamf6 was prominently observed in tumor-reactive CD8+T cells in the lungs, in line with the notion that Slamf6 functions as a representative molecule for Texp cells in chronic infections or tumor models [12] (supplemental figure S1c). Based on these findings, we hypothesized that activated CD8+ T-cells may play a crucial role in the regulation of metastasis. To further investigate this phenomenon, we isolated lung-derived tumor-specific effector-like Tex cells, based on the expression of the surface molecule Slamf6−CX3CR1+, and subsequently re-transferred them into Cd8−/− mice bearing metastases for therapeutic purposes. In addition, we concurrently processed Texp cells (identified as Slamf6+Tim3−) and terminal Tex cells (characterized as Slamf6−CD101+). Our findings revealed that the transfer of tumor-specific effector-like Tex cells did not confer any survival advantage; however, the administration of Texp cells resulted in remarkable remission of metastasis (Fig. 1d). Moreover, the infusion of precursor cells at different time points yielded distinct advantages in terms of the overall survival duration for mice with metastases (Fig. 1e). Collectively, these findings support the essential role of Texp cells derived from metastatic tissues in protecting against the invasion of metastatic lesions.
3.2 CXCR6+ subsets are present in Texp cellsSubsequently, we examined the distinctive characteristics exhibited by tumor-specific Texp cells during metastasis. Chemokine receptors play a crucial role in mediating CD8+ T-cell responses against tumors [20,21,22,23]; therefore, we examined their relative expression in metastatic tissues. A total of 5 × 105 CD45.1+OT-I cells were transferred to mice on day − 2, after the intravenous administration of B16-OVA cells, lung tissues were harvested and subjected to CD44+OT-I cell analysis on the indicated days. To eliminate potential contamination from circulating CD8+ T cells in the peripheral blood, mice were intravenously administered anti-CD8 (APC-cy7 channel) prior to sacrifice, enabling the identification of lung-resident CD8+ T cells that lacked APC-cy7 staining. Notably, we observed a subset of Texp cells with pronounced upregulation of CXCR6 but lacking obvious expression of CCR2, CXCR3, CXCR5, or CX3CR1; however, this expression gradually declined at subsequent time points (Fig. 2a) (supplemental figure S2a). Moreover, a comparable expression pattern was observed in endogenous tumor-reactive CD8+ T cells (identified as OVA257 − 264 peptide (SIINFEKL) H-2Kb tetramer-positive) within metastatic mice (supplemental figure S2b). However, the number of CXCR6+ Texp cells within the subcutaneous tumors was limited (supplemental figure S2c).
Fig. 2
Expression pattern of CXCR6 + Texp cells (a). C57BL/6J mice were intravenously(i.v) injected with 5 × 105 of CD45.1+ OT-I cells prior to B16-OVA cells inoculation (n = 5 per group), on day 8, 16 and 26, lung-derived TILs were analyzed by flow cytometry. Representative flow cytometry staining (left) and summary (right) of Slamf6/CXCR6 expression on OT-I cells (gated from live CD45.1+CD44+) were calculated. (b). CD45.1+ OT-I cells (5 × 105) were adoptively transferred into C57BL/6J mice (n = 4 per group) on day − 2, followed by 3 × 105 of B16-OVA cells inoculation. From day 3 to day 7 post transfer, mice were intraperitoneally (i.p) treated with FTY720 at a dose of 2.5 mg/kg every 2 days. Lung-derived TILs were subjected to flow cytometry, the absolute number of Slamf6+CXCR6+ OT-I cells were measured. In (a, b), statistical comparisons were performed using an unpaired t-test, ns, no significance, *p < 0.05, **p < 0.01, ***p < 0.001. Data are shown as the mean ± s.e.m. and representative of two independent experiments
Recent studies have revealed the crucial role of tumor-draining lymph nodes in facilitating CD8+ T cell infiltration into the solid tumor microenvironment [24,25,26]. We aimed to investigate whether tissue-derived CXCR6+ Texp cells could migrate from lung-draining mediastinal lymph nodes (MLNs). Given that surgical resection of MLNs is challenging, we opted for a pharmacological intervention using FTY720 to effectively impede cell migration from MLNs to the peripheral blood. Mice were adoptively transferred to OT-I cells, followed by inoculation with B16-OVA cells. Intraperitoneal administration of FTY720 was performed three times. Notably, administration of FTY720 significantly attenuated the absolute count of CXCR6+ Texp cells in the lungs (Fig. 2b), highlighting the pivotal role of MLNs as a source of Texp cells during metastasis. Moreover, we investigated whether tumor antigen burden accounted for the generation of the CXCR6+ population. Our findings revealed an elevated proportion of these cells in response to high tumor cell engraftment in lung tissues (supplemental figure S2d), implying an antigen-driven mechanism underlying CXCR6+ Texp cell generation.
3.3 CXCR6 + Texp cells exhibit limited cellular memory attributesTexp cells have been reported to exhibit nonclassical memory cell characteristics within the tumor microenvironment [27, 28]. A total of 5 × 105 CD45.1+ OT-I cells were adoptively transferred into mice prior to inoculation with B16-OVA cells. Subsequently, the lung tissue was harvested and expanded Texp cells were subjected to flow cytometric analysis. We observed that CXCR6+ subsets exhibited significantly lower levels of memory-associated markers CD62L, IL2Rβ, or IL7Rα in comparison to CXCR6− cells on day 12 post-cell transfer (Fig. 3a). In addition, we investigated the expression profiles of inhibitory markers, including PD-1, Tim3, Lag3, CD39 and Tox, in tumor-specific CXCR6+ Texp cells, and terminal Tex cells. These distinctive marker expressions were observed in nearly all terminal Tex cells but were absent in CXCR6+ cells (Fig. 3b).
Fig. 3
The molecular characteristics associated with memory of Texp cells. C57BL/6J mice were adoptively transferred with 5 × 105 of splenic CD45.1+ OT-I cells on day − 2, followed by intravenous injection with B16-OVA cells. On day 12 post transfer, lung-derived TILs were isolated and subjected to flow cytometry. (a). The representative staining and quantification of IL2Rβ, IL7Rα or CD62L expression on CXCR6+ and CXCR6− Texp subsets; (b). Flow cytometry analysis of inhibitor factors PD-1, Tim3, Lag3, CD39, Tox expression on CXCR6+ Texp cells and terminal Tex cells. Cells were gated from live CD45.1+CD44+ cells. The experiments were performed at least twice with similar results (n = 5 per group). c The schematic of experimental design (left) and the flow cytometry analysis (right) of in vivo co-transfer assay. The Texp cells utilized in the flow cytometry analysis originated from the lung tissue of the host mouse. d Heatmap of different gene expressions of antigen-specific OT-I cells by using q-PCR. Mice were adoptively transferred with naïve OT-I cells on day − 1 and were intranasally infected with influenza A/HKx31 containing OVA peptide at 300 EID50; then lung tissues were harvested and tissue-resident memory CD8+ T cell (Trm) cells were isolated on day 40 post infection. Lung-infiltrated Texp cells were processed as above described. In (a-b), statistical comparisons were performed using an unpaired t-test, **p < 0.01, ***p < 0.001. Data are shown as the mean ± s.e.m. and representative of two independent experiments
The long-term persistence of memory CD8+ T cells orchestrated by homeostatic proliferation represents a fundamental mechanism that has been demonstrated to affect the establishment of anti-reinfection or anti-tumor immunity [29, 30]. To assess the long-term potential, metastatic lung tissues were harvested and Texp cells were isolated by flow cytometry. Subsequently, CXCR6+ cells (CD45.1+CD45.2+) and CXCR6− cells (CD45.1+) were co-transferred into recipient mice with a CD45.2+ background at a 1:1 ratio. After resting for 14 days, the samples displayed a significant decrease in the proportion of CXCR6+ cells (Fig. 3c), indicating that tumor-reactive CXCR6− Texp cells exhibit greater longevity and sustainability, whereas this function is diminished in CXCR6+ cells. In addition, to elucidate the transcriptional profiles, we employed flow cytometry to sort lung-infiltrated Texp cells from mice with metastasis and Trm cells (identified by CD69+CD103+) from mice infected with the influenza virus. Subsequently, genotypic expression was examined by real-time qPCR analysis. Our findings revealed that CXCR6− Texp cells exhibited comparable levels of gene expression encoding signature transcription factors associated with Trm cells, such as Runx3, Tcf7, Sell, whereas CXCR6+ cells showed a weak expression pattern (Fig. 3d).
We investigated the essentiality of IL-15 and TGFβ, two important cytokines for maintaining memory CD8+ T cells, in regulating the functionality of CXCR6+ Texp cells during metastasis. Prior to melanoma cell inoculation, naïve OT-I cells were adoptively transferred into mice, followed by the administration of blocking antibodies against IL-15 and TGFβ, administered three times. Lung-derived Texp cells were analyzed using flow cytometry. Treatment with both anti-IL15 and anti-TGFβ resulted in a significant reduction in the absolute number of CXCR6− Texp cells, however, the number of CXCR6+ Texp cells was scarcely affected (supplemental figure S3a). Collectively, these findings suggest that tumor-specific CXCR6− Texp cells may possess a memory-like imprint characterized by specific phenotypes, transcription factors, and self-renewal capacity; however, these characteristics are predominantly diminished in the context of metastasis among CXCR6+ Texp cells.
3.4 Generation of effector-like Tex cells predominantly relied on CXCR6+ Texp cells in metastasisWe investigated the role of Texp cells in metastasis. Approximately 5 × 105 OT-I cells were adoptively transferred into mice, followed by the intravenous inoculation of B16-OVA cells. On day 8 post-cell transfer, the lungs were harvested, and Texp cells were isolated using flow cytometry. After stimulation with the OVA257 − 264 peptide in vitro, along with anti-CD28 and IL-2 for 48 h, CXCR6+ Texp cells exhibited a robust proliferative response (supplemental figure S4a), indicating their potential for rapid cell differentiation. Purified CXCR6+ Texp cells were re-transferred into metastasis-matched mice, where a fraction (20–40%) of donor cells differentiated into CX3CR1-expressing effector-like Tex cells at the initial time point examined in the lungs; however, only a few terminal Tex counterparts were detected (Fig. 4a). Subsequent analysis revealed a significant increase in the frequency of terminal Tex subsets and a gradual decrease in the number of newly formed effector-like Tex cells. These findings suggest that CXCR6+ Texp cells can promptly replenish the early population of effector-like Tex cells during metastatic progression.
Fig. 4
CXCR6 + Texp cells served as a crucial source of effector-like Tex cells (a). Representative flow cytometry staining (left) and summary (right) of differentiation pattern on effector-like and terminal Tex subsets at different time points. A total of 5 × 105 of CD45.1+OT-I cells were adoptively transferred into C57BL/6J mice subsequently intravenous injection with B16-OVA cells. On day 8 post tumor cell inoculation, lung tissues were harvested, and CXCR6+Texp cells were purified and re-transfer into metastasis-matched mice. b The flow cytometry analyses of the percentages of TCF1−CX3CR1+ OT-I cells. CD45.1+CXCR6− and CD45.1/2+CXCR6+ Texp cells were sorted from lungs of metastatic mice on day 8 and were co-transfer in a 1:1 ration (a total of 6 × 105) into metastasis-matched mice inoculated with B16-OVA cells (n = 3 per group) (c). The schematics of experimental design of in vivo co-transfer assay. Naïve Cxcr6 deficient Texp cells were isolated from CD45.1+ background mice, and equally mixed with CD45.1+CD45.2+ normal Texp cells to co-transfer into C57BL/6J mice. Statistical comparisons were performed using an unpaired t-test, **p < 0.01. Data are shown as the mean ± s.e.m. with 6 mice per group and representative of two independent experiments
To validate the replenishment of effector-like Tex subsets in metastatic tissues, we performed an in vivo cell co-transfer assay. Lung-derived CXCR6+/CXCR6− Texp cells from different donors were sorted by flow cytometry and co-transferred in a 1:1 ratio into recipient mice with matched metastases. On day 6 post-cell transfer, more than half of the CXCR6+ cells underwent differentiation into effector-like Tex counterparts, whereas most donor CXCR6− cells exhibited sustained Slamf6 expression (Fig. 4b). In addition, CD45.1+Cxcr6−/− and CD45.1/2+Cxcr6+/+ OT-I cells were co-transferred in equal numbers into naïve mice, followed by inoculation with B16-OVA cells. Metastatic lungs were harvested, and effector-like Tex cells were analyzed by flow cytometry on day 8 post-melanoma cell inoculation. A significant reduction in the frequency of expanded effector-like Tex cells was observed in donors with CXCR6-deficient cells compared to that in their normal counterparts (Fig. 4c). Collectively, these findings suggest that CXCR6+ Texp cells play a crucial role in generating functional CD8+ T cells against tumors within the metastatic microenvironment, while delayed differentiation was observed in their CXCR6− counterparts.
3.5 CXCR6+ Texp cells exhibit potent control over lung metastasisTo investigate the role of antigen-specific Texp cells in metastasis control, approximately 1 × 106 cells were isolated from metastatic lungs and subsequently transferred into pre-established Cd8−/− mice with metastases (mice with homogeneous levels of lucifer fluorescence were incorporated into the treatment group) (Fig. 5a). A substantial proportion of donor cells exhibited lung homing following the transfer (supplemental figure S5a). The number of metastatic foci was quantified on day 16 post-adoptive transfer. Mice receiving CXCR6+ Texp cells showed a significant reduction in the number of metastatic foci compared to the other two groups (Fig. 5a). In addition, we evaluated the survival duration of these recipient mice and observed that nearly half of them exhibited a prolonged lifespan of up to 60 days following the transfer of CXCR6+ Texp cells, which was significantly longer than that of the control mice (Fig. 5b). Furthermore, we implemented a combined therapeutic approach involving cellular adoptive transfer and cyclophosphamide (CTX) administration in pre-established lung metastasis-bearing mice. We observed that treatment with CTX alone extended the lifespan of mice; however, when combined with the adoptive transfer of CXCR6+ Texp cells, it significantly enhanced therapeutic efficacy against metastasis (Fig. 5c). However, the effectiveness of adoptive transfer of CXCR6+ Texp cells appears to be limited in the context of targeting subcutaneous tumors (supplemental figure S5b). Collectively, these findings highlight the potential value of Texp cell transfer as a promising therapeutic strategy for effectively managing metastasis.
Fig. 5
CXCR6 + Texp cells effectively mitigated lung metastasis (a-d). The schematics of experimental design of in vivo cell-transfer immunotherapy assay and survival curves of control or treated mice. No treatment mice was used as control. Log rank test of survival data was used. Approximately 1 × 106 of lung-derived CXCR6−/CXCR6+ Texp cells were sorted from metastatic donor mice on day 12 as above described. Cells were adoptively transferred into pre-established metastasis-bearing Cd8−/− mice 4 days post-surgery, respectively. The representative images of lung nodules (a) or survival curve (b) of mice were measured. Red line versus gray line, p = 0.068; Blue line versus Red line, p < 0.01. c The recipient mice were administrated with cyclophosphamide (CTX, sigma) drug at a dose of 350 mg/kg i.p. twice prior to CXCR6+ Texp cells transfer (1 × 106). Green line versus blue line, p < 0.05; Red line versus blue line, p < 0.01; Red line versus green line, p < 0.05. d A total of 1 × 106 of CXCR6−/CXCR6+ Texp cells were adoptively transferred into C57BL/6J mice subsequently subcutaneous footpad inoculation with B16-OVA cells (1 × 106). Blue line versus gray line, p < 0.01; Red line versus blue line, p < 0.01.In (a), statistical comparisons were performed using an unpaired t-test, *p < 0.05, **p < 0.01. Data are shown as the mean ± s.e.m. with 4–7 mice per group and representative of two independent experiments
To investigate the potential impact of CXCR6+ subset deficiency on the suppressive capacity of CD8+ T cells in metastasis, we established a bone marrow chimeric mouse model by combining bone cells derived from C57 donors and Cxcr6−/− donors or Cd8−/− donors and Cxcr6−/− donors, followed by injection of cell mixtures into sub-lethally irradiated Cd8−/− recipients. After 10-week remodeling, the recipient mice were challenged with B16-OVA cells. The results revealed that the absence of CXCR6+ subsets failed to confer effective protection against metastasis in mice (supplemental figure S5c).
We performed a therapeutic experiment wherein purified lung-derived Texp cells were adoptively transferred into Cd8−/− recipient mice on day − 2 after B16-OVA inoculation (Fig. 5d). Transfer of CXCR6− cells conferred substantial protection against lung metastasis, whereas mice receiving CXCR6+ cell transfer exhibited limited survival. These results indicate that CXCR6+ Texp cells have the capacity to control lung metastasis, while CXCR6− cells exhibit preventive effects against it.
3.6 CXCR6+ Texp cells exert metastasis control through diverse mechanismsBased on our findings, adoptive transfer of tumor-specific CXCR6+ Texp cells has the potential to mitigate metastasis. We postulated that CXCR6+ Texp cells exert direct cytotoxic effects against tumor cells. To test this hypothesis, naïve CD45.1+ OT-I cells were adoptively transferred into C57 mice, followed by administration of B16-OVA cells. Lung tissues were harvested and expanded CD45.1+ OT-I cells were analyzed using flow cytometry. We observed that CXCR6+ Texp cells exhibited a high expression of the activation-associated marker KLRG1, whereas minimal expression was detected on CXCR6− cells at day 12 post-cell transfer (Fig. 6a). Moreover, transfer of CXCR6+ Texp cells showed augmented cytotoxicity by upregulating the production of cytokines including IFN-γ, TNFα, CD107a/b, and GzmB, which are associated with cytolytic function (Fig. 6b). To confirm the cytotoxicity of CXCR6+ Texp cells against tumor cells, we performed an in vitro killing assay (supplemental figure S6a). Lung-derived CXCR6+ Texp cells were isolated from metastasis-bearing mice and co-cultured with CFSE-labeled target cells for 12 h, followed by flow cytometry. The attenuation of the fluorescent signal implies that the target cell has been effectively eliminated. Notably, these cells exhibited potent direct cytotoxicity as evidenced by a significant reduction in the target cell population (Fig. 6c). Furthermore, CXCR6+ Texp cells were activated, sorted, and co-cultured with CFSE-labeled B16-OVA cells before being re-transferred into Cd8−/− recipient mice. Naïve OT-I cells were used as controls, and apoptosis of melanoma cells in the lungs was evaluated using flow cytometry. Notably, the transfer of CXCR6+ Texp cells significantly increased the percentage of 7-AAD+ annexin V+ melanoma cells in vivo (supplemental figure S6b).
Fig. 6
CXCR6+ Texp cells exerted direct cytotoxic effects on tumor cells. (a-b) Approximately 5 × 105 of splenic CD45.1+ OT-I cells were transferred into C57BL/6J mice (n = 4 per group) subsequently intravenous injection with 3 × 105 of B16-OVA cells. On day 12 post transfer, lungs were harvested and TILs were processed to flow cytometry. Flow cytometry analyzing KLRG1 expression (a), or granzyme B(Gzmb), CD107a/b, tumor necrosis factor-α (TNFα) and IFNγ expressions (b), on CXCR6−/CXCR6+ Texp cells. (c). Flow cytometry data of the in vitro killing capacity assay of CXCR6+ Texp cells. The details as shown in methods and supplemental figure S6a. (d). Flow cytometry analyzing the frequency of IFNγ+CD8+ T cells after in vitro stimulation with OVA257 − 264 peptide along with PMA, ionomycin and GolgiPlug/stop for 5 hours. The assay design as shown in supplemental figure S6d. In (b, d) statistical comparisons were performed using an unpaired t-test, *p < 0.05, ***p < 0.001. Data are shown as the mean ± s.e.m. and representative of two independent experiments
The absolute number of dendritic cell (DC) expressing CD11c and MHCII, which play a crucial role in antigen recognition by CD8+ T cells within the tumor microenvironment, was significantly higher in the lungs of recipient mice following the transfer of CXCR6+ Texp cells than in the control group (supplemental figure S6c). Previous study have demonstrated the substantial presence of bystander CD8+ T cells within tumor infiltrates [31]. Isolated CXCR6+ Texp cells (CD45.1+) were re-transferred into naïve C57 recipients, followed by the administration of B16-OVA cells. On day 16 post-transfer, endogenous CD45.2+CD8+ T cells were sorted and cultured with OVA257 − 264 peptide in vitro. Upon stimulation, a significant increase in the proportion of IFN-gamma-producing CD8+ T cells was observed among mice that received a transfer of CXCR6+ Texp cells, indicating the augmented presence of tumor-reactive CD8+ T cells (Fig. 6d) (supplemental figure S6d).
3.7 CXCR6 + Texp cells can respond to ICBCancer immunotherapy involving ICB with monoclonal antibodies targeting PD-1/PD-L1 has demonstrated the ability to restore the impaired function of cytotoxic effector CD8+ T cells, resulting in tumor regression and prolonged patient survival [8, 32, 33]. However, the subsets of CD8+ T cells that respond to ICB in tumors remain elusive. Based on our findings, a minority of tumor-specific CXCR6+ Texp cells exhibit PD1 protein expression during metastasis, indicating their potential responsiveness to ICB. To test this hypothesis, we isolated lung-derived CXCR6+ Texp cells from metastasis-bearing mice and cultured them with an anti-PD1 antibody in vitro along with anti-CD3 and anti-CD28 antibodies. After treatment with the anti-PD1 antibody, there was a two-fold increase in the absolute number of Texp cells, indicating that antigen-specific CXCR6+ Texp cells responded to the PD1 blockade (Fig. 7a). To confirm this observation, we transferred isolated CXCR6+ Texp cells into pre-established metastatic mice and administered three doses of anti-PD1, while monitoring cell proliferation using BrdU dye. A significant increase in donor-derived BrdU+ cells was observed following anti-PD1 treatment (Fig. 7b). However, endogenous tumor-specific CD8+ T cells exhibited a slight upregulation of BrdU expression (supplemental figure S7a). Collectively, these findings demonstrate the capacity of CXCR6+ Texp cells to respond to ICB and potentially enhance the therapeutic efficacy of ACT.
Fig. 7
CXCR6 + Texp cells could respond to immune checkpoint blockade (a). Flow cytometry analyzing absolute number of CXCR6+Texp cells which were isolated from metastatic mice on day 8 post cell transfer, followed by in vitro stimulation with IL2 and anti-PD1 mAb in CD3/CD28 mAbs-coated culture plates for 72 h. (b). The assay design of in vivo anti-PD1 mAb treatment and the representative flow cytometry staining of Brdu on CD44+CD45.1+CXCR6+Texp cells in metastasis-matched mice. (c). Approximately 1 × 106 of CXCR6+Texp cells as described in (a) were adoptively transferred into metastasis-bearing Cd8−/− mice. Recipient mice were administrated with anti-PD-1 or control antibody at dose of 6 mg/kg on 5, 8, 11 days post cell transfer. No treatment mice was used as control. Survival curves of mice were calculated. Log rank test was used. Blue line versus gray line, p < 0.01; Red line versus blue line, p = 0.037
To investigate the potential synergistic effect of the adoptive transfer of tumor-specific CXCR6+ Texp cells in combination with ICB, we adoptively transferred these cells into metastasis-bearing Cd8−/− mice prior to the administration of anti-PD1 antibodies. Combination therapy involving CXCR6+ Texp cells and anti-PD1 blockade demonstrated superior therapeutic efficacy compared to individual treatments in our study (Fig. 7c).
Comments (0)