
Remember me


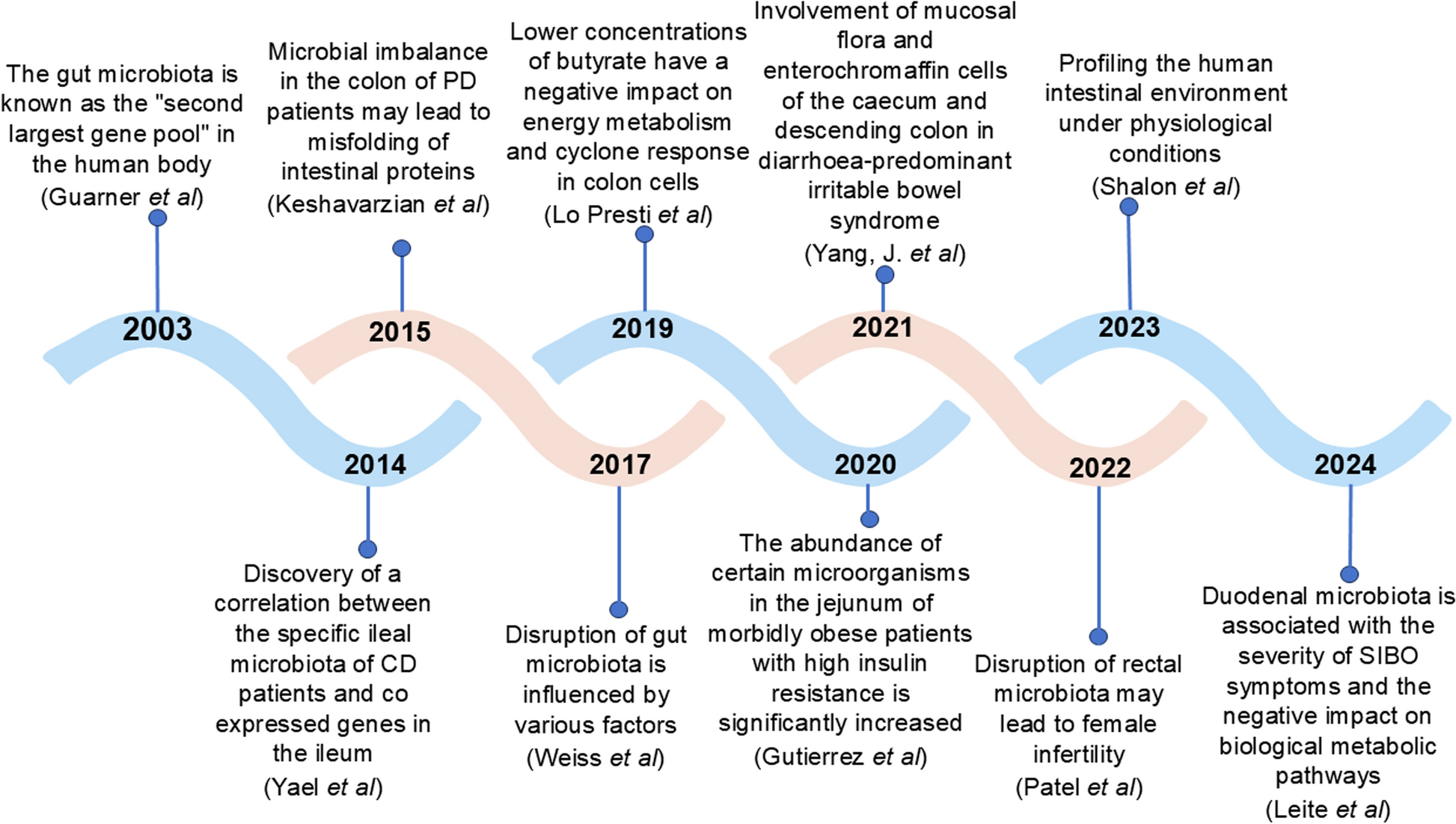





GM is intricately linked to numerous physiological and pathological processes in humans. Alterations in its composition and distribution significantly influence the progression of various clinical diseases. However, most research has primarily utilized fecal samples to explore the relationship between diseases and GM, with limited focus on potential differences in microbiota across different anatomical sites [45]. Recent studies have revealed substantial disparities between the microbiota and its metabolites in the actual gut environment compared to those in fecal samples, indicating that fecal microbiota and metabolites are not fully representative of the microbial composition and metabolic status of the entire intestine [46, 47]. The composition of the microbiota varies across different intestinal segments, including duodenum, jejunum, ileum, cecum, colon and rectum, and their variations exert distinct effects on disease procession (Table 2). Recently, some studies have begun exploring the relationship between diseases and microbial changes in specific intestinal segments, which could facilitate the precise regulation of the microbiota in targeted intestinal segments for microbiome-based therapeutics in related diseases.
Table 2 Alterations of microbes in different intestinal segment affecting different diseases and their potential mechanismsDuodenal microbiota and diseaseFunctional dyspepsia (FD) is a clinical syndrome characterized by epigastric pain or burning, postprandial fullness, or early satiety lasting for at least 6 months [48]. Studies have shown that the duodenal microbiota of FD patients undergoes significant changes in microbial biomass and diversity. For example, Shanahan [49] found that the relative abundance of Firmicutes, Bacteroidetes and Fusobacteria in the duodenal MAM of 56 FD patients correlated with the severity of disease symptoms. At the genus level, the relative abundance of Rothia, Clostridium, Haemophilus, and Actinobacillus significantly increased [50], while the abundance of Streptococcus, Prevotella, and Veillonella spp. was negatively correlated with gastric emptying time [51]. Additionally, the duodenal mucosa of FD patients exhibits low-grade inflammation and destruction of the mucosal barrier, with increased mucosal permeability correlating with the severity of inflammation, but whether the microbiota is involved in this process is not clear [52]. These findings suggest that duodenal inflammation in FD patients may be driven by dysbiosis, although the exact pathogenesis—such as how duodenal microbiota contributes to intestinal inflammation and gastrointestinal symptoms—requires further investigation.
Celiac disease (CeD) is an inflammatory disease of the small intestinal mucosa triggered by intolerance to gluten containing cereals [53]. In CeD patients, the duodenal microbiota shows an increase in Proteobacteria and a decrease in Firmicutes and Bacteroidetes. At the genus level, there is an increase in Enterobacter, Streptococcus and Escherichia, alongside a decrease in Akkermansia, Clostridium, and Bacillus [54,55,56]. Moreover, the abundance of Neisseria correlates with the degree of intestinal villus atrophy. Notably, Neisseria flavescens can evade lysosomal degradation in Caco-2 cells and induce inflammatory responses in dendritic cells and isolated mucosal explants [57]. In addition, Pseudomonas aeruginosa participates in gluten hydrolysis, altering its immunogenicity and activating the innate immune pathway associated with CeD [58]. These findings indicate that the duodenal microbiota may stimulate the intestinal immune system and induce a variety of inflammatory signals in CeD by degrading gluten into immunogenic peptides. However, the specific mechanism remains to be fully elucidated and verified.
Recent research has identified the impact of duodenal microbiota on the pathogenesis of pancreatitis and pancreatic cancer. In the duodenal MAM of patients with acute pancreatitis (AP), there is a significant increase in the relative abundance of Pseudomonas, Ruminococcaceae, and Pediococcus, along with enriched functional pathway related to the endocrine system, glycerolipid metabolism, and dioxin degradation [59]. Additionally, in patients with pancreatic cancer, the duodenal MAM shows higher relative abundances of Acinetobacter, Aquabacterium, Oceanobacillus, Rahnella, Massilia, Delftia, Deinococcus, and Sphingobium were higher, while Bacteroides abundance decreases, possibly linked to increased duodenal mucosal inflammation [60, 61]. Kohi [62] also found that in patients with pancreatic ductal adenocarcinoma (PDAC), the duodenal microbiota was dominated by Enterococcaceae, Lactobacillaceae, and Bifidobacteriaceae dominated the duodenal microbiota, and Fusobacterium, Rothia, and Neisseria significantly enriched in short-term PDAC survivors. These findings suggest that alterations in the abundance of specific duodenal microbiota may induce pancreatic inflammation and affect pancreatic cancer progression by modulating related human metabolic pathways. Consequently, the role of duodenal microbiota in pancreatic cancer needs to be further investigated to clarify its potential in better monitoring patients for pancreatic cancer risk.
Jejunal microbiota and diseaseSmall intestinal bacterial overgrowth (SIBO) has traditionally been diagnosed using small bowel aspirate, considered the ‘gold standard’, with a threshold of 105 CFU/mL in jejunal aspirate and 103 CFU/mL in duodenal aspirate. However, glucose and lactulose breath tests are now commonly used in clinical practice for SIBO diagnosis: defined by an increase in H2 concentration of 20 parts per million (ppm) from baseline within 90 min and an increase in CH4 levels ≥ 10 ppm at any time [63]. Common clinical symptoms of SIBO include abdominal pain, bloating, diarrhea, flatulence and dyspepsia [64], which may be associated with translocation and overgrowth of jejunal microbiota. For example, Pistiki [65] isolated approximately 170 species of aerobes from the jejunum of 117 SIBO patients, mainly including Klebsiella, Enterobacter, Enterococcus faecalis, Escherichia coli, Pseudomonas aeruginosa, and Staphylococcus aureus. Additionally, small bowel surgery, radiation or radiotherapy can all lead to the occurrence of bacterial translocation [66], due to factors such as inhibited intestinal peristalsis, abnormal intestinal structure or reduced gastric acid secretion, providing opportunities for bacterial overgrowth. This overgrowth results in the excessive production of acid and gas, causing abdominal pain and bloating [67]. Furthermore, the growth of certain translocated microbiota in the jejunum can lead to metabolic disorders. For instance, Lactobacillus can produce bile salt hydrolase or lipase, leading to the depolymerization and premature reabsorption of conjugated bile acids in the jejunum instead of the ileum. This interference with enterohepatic circulation results in fat malabsorption [28, 29]. Meanwhile, free bile acids, which are toxic to the intestinal mucosa, can lead to bile acid diarrhea and further malabsorption [66]. In summary, the translocation and overgrowth of jejunal microbiota are closely related to the development of SIBO, and changes in the abundance of different strains trigger various gastrointestinal symptoms. However, the specific roles of many strains in the jejunal microbiota in SIBO require further studies.
The jejunal microbiota has also been implicated in diseases related to dysregulation of glucose homeostasis, such as obesity or diabetes. Carolina [68] observed that in the jejunal MAM of morbidly obese patients with high insulin resistance, the abundance of Proteobacteria, Fusobacteria, and Bacteroidetes significantly increased. Metformin treatment notably upregulated the relative abundance of Halomonadaceae and markedly slowed disease progression, suggesting that Halomonadaceae in the jejunal microbiota plays a role in morbidly obese patients with high insulin resistance. In addition, Jayashree [69] found that intestinal permeability (IP) was significantly higher in patients with type 2 diabetes mellitus (T2DM) compared to healthy adults, a finding also observed in T2DM rat models compared to normal rats [70]. The level of IP was significantly negatively correlated with the relative abundance of Bacteroides fragilis [71], indicating that a decrease in Bacteroides fragilis may be a critical factor contributing to jejunal barrier damage. Furthermore, the jejunal microbiota of the T2DM rat model exhibited a significant increase in Proteobacteria, Bacteroidetes, Acidobacteria, Gemmatimonadetes, and TM7. At the genus level, Shigella, Alcaligenes, Bacteroides, Allobaculum, and Clostridiaceae Clostridium were also significantly reduced [71]. Current research suggests that glucose degradation may be the main pathway through which the jejunal microbiota regulates host metabolism to influence obesity and T2DM [72]. In general, disturbances in the jejunal microbiota may lead to damage of the jejunal barrier, contributing to the pathogenesis of obesity or diabetes. However, the association between jejunal microbiota and glucose metabolism requires further exploration and clarification.
In summary, each intestinal segment harbors unique physiological functions and distinct microbiota compositions that collectively contribute to overall gut homeostasis and host health. Understanding the specific interactions between microbiota and their respective intestinal environments is essential for elucidating their roles in various physiological and pathological processes. This knowledge lays the foundation for developing targeted microbiome-based diagnostic and therapeutic strategies for a wide range of diseases.
Ileal microbiota and diseaseCrohn's disease (CD) is a chronic inflammatory bowel disease with an unclear etiology. Studies have demonstrated significant differences in the composition of the ileal microbiota across various stages of CD, suggesting that interactions between the ileal microbiota and the host play a crucial role in the disease's pathogenesis [73]. For example, the relative abundance of Bacteroides is higher during the active stage of CD, whereas Streptococcus, Veillonella, and Lactococcus were higher in the remission. These bacteria are primarily involved in amino acid and carbohydrate transport and metabolism [74]. Yael [75] found that specific ileal microbiota of CD patients are associated with the co-expression genes APOA1 and DUOX2 in the ileum. Specifically, up-regulation of DUOX2 gene correlates with an increase in Proteobacteria, while downregulation of APOA1 is associated with a decrease in Firmicutes and Bacteroidetes. These changes promote oxidative stress and Th1 cell polarisation, exacerbating intestinal mucosal damage in CD patients. Additionally, recurrence of CD following ileal resection is common and may be related to a reduction in the alpha diversity of the ileal mucosa-associated microbiota (MAM), an increase in certain Proteobacteria members, and a decrease in members of the Lachnospiraceae and the Ruminococcaceae families within the Firmicutes phylum [76, 77]. These microbial alterations may serve as more specific predictors of CD recurrence risk compared to general clinical factors. Overall, the distinct microbial and genetic profiles of the ileum identify it as a primary inductive site for all forms of CD, potentially aiding in the diagnosis of different disease stages, therapeutic selection, and prognosis evaluation.
The ileal microbiota also influences the local immune microenvironment and participates in immune surveillance of colon cancer (CC). Research indicates that ileal microbiota affects the therapeutic efficacy of oxaliplatin in CC patients by modulating tumor immunosurveillance. Marion [78] observed that Bacteroides fragilis was the only enriched ileal microbe in CC patients with a favorable prognosis, whereas Paraprevotella clara predominated in those with a poor prognosis. In the CC mouse model, mice supplemented with Bacteroides fragilis exhibited higher levels of tumor-infiltrating lymphocytes (TILs) and lower levels of CD45 cells in the ileum compared to those supplemented with Paraprevotella clara [78]. These findings align with previous studies showing that B. fragilis induces dendritic cells to release IL-1β and IL-12, increasing the number of follicular helper T (Tfh) cells, while Fusobacterium nucleatum promotes Th17 cell accumulation in tumor-draining lymph nodes, enhancing the immune response [79, 80]. Thus, specific ileal microbiota, such as B. fragilis and F. nucleatum, may be involved in the pathogenesis and therapeutic response of CC by modulating the ileal immune response.
Recent studies have also linked the ileal microbiota to the host's circadian rhythm. For example, Ana Carolina [81] found that in Cry1−/−Cry2−/−rhythm gene knockout mouse model, eating and sleeping rhythms were significantly disrupted, leading to the complete loss of microbial cycling in the ileum Additionally, Erysipelotrichaceae and Lactobacillaceae were significantly increased in the ileal microbiota. This disruption mirrors findings from other studies where knockout of clock genes such as Bmal and Per led to disturbances in the microbial circadian rhythm [82]. Furthermore, the ileal microbiota appears to be linked to the circadian rhythm of the host liver. Disruption of the microbiota's circadian dynamics affects the liver transcriptome, leading to non-rhythmic expression of genes involved in chromatin and nucleosome assembly, as well as the metabolism of amino acids, polysaccharides, lipids, and steroids [83, 84]. Consequently, the host's molecular biological clock is intertwined with the diurnal dynamics of the ileal microbiota. Interference with the host's biological clock may disrupt the circadian rhythm of the ileal microbiota, subsequently affecting hepatic metabolic rhythms.
Cecal microbiota and diseaseThe cecal microbiota is associated with diseases related to abnormal lipid metabolism. For example, Yuan [85] found a significant increase in the relative abundance of Cyanobacteria, Rhodospirillaceae, Olsenella, Sporobacter, Anaerovorax, Insolitispirillum, and Blautia in the cecal microbiota of mice with furan-induced liver injury. Notably, Cyanobacteria produce Microcystins-LR, which can disrupt redox balance and impair liver function in mice [86]. Therefore, Cyanobacteria may serve as a potential marker for liver injury, providing new ideas for subsequent research on liver injury. Additionally, Xueliang [87] found that in the cecal microbiota of mice with non-alcoholic fatty liver disease(NAFLD), the relative abundance of Bacteroidetes was low, while Firmicutes was high, resulting in an increased Firmicutes / Bacteroidetes ratio. Microbes positively correlated with lipid content, such as Paraprevotella, Clostridium Xl, and Barnesiella, were increased, whereas those negatively correlated with fat accumulation, such as Akkermansia, Alloprevotella, and Parabacteroides were decreased. Additionally, Veillonella and Olsonella were positively correlated with serum ALT and ALP levels [87]. In rat models of high-fat diet (HFD)-induced hyperlipidemia, there was an increase in Firmicutes and a decrease in Euryarchaeota, Actinobacteria, and Tenericutes within the cecal microbiota [88]. Furthermore, there was also a significant decrease in Corynebacterium_1, Nosocomiicoccus, and Jeotgalicoccus, alongside an increase in Turicibacter and Bifidobacterium. Metabolites such as linoleic acid and sphingosine were also significantly elevated [88]. To sum up, the cecal microbiota may play a role in regulating diseases like hyperlipidemia and NAFLD through disruptions in lipid metabolism. However, the specific mechanisms remain to be further explored.
The Cecal microbiota is also implicated in other diseases. For example, in the cecal samples from patients with diarrhea-predominant irritable bowel syndrome (IBS-D), the relative abundance of Ruminococcus torques increased, and the relative abundance of Ruminococcus torques decreased [89]. This aligns with studies showing a positive correlation between Ruminococcus torques and enterochromaffin cells, IBS-SSS, degree of abdominal pain, frequency of abdominal pain, and frequency of defecation [90, 91]. Additionally, a significant decrease in short-chain fatty acids that enhance macrophage function to control inflammation, was found in the cecal contents of mice with Klebsiella pneumoniae-Induced Pneumosepsis [92]. This suggests that bacterial pneumonia can dramatically alter the GM and metabolites, leading to a dysregulated immune response to the lung infections. In conclusion, while the cecal microbiota has been preliminarily linked to various diseases, the specific roles and mechanisms require further investigation, including validation in clinical translational studies.
Colonic microbiota and diseaseThe colonic microbiota is relevant with several diseases, such as inflammatory bowel disease (IBD), irritable bowel syndrome (IBS), and colorectal cancer (CRC). IBD is a chronic and recurrent inflammatory disease of the digestive system mediated by immune response, including CD and ulcerative colitis (UC) [93]. In IBD patients, there is a significant increase in Enterobacteriaceae and a notable decrease in butyrate-producing microbes such as Faecalibacterium prausnitzii and members of the Ruminococcaceae family. This results in lower butyrate concentrations, negatively impacting energy metabolism and inflammatory responses in colon cells [94, 95].
IBS is a functional gastrointestinal disorder with symptoms that include abdominal pain and changes in stool or frequency [96]. In the colonic MAM of IBS patients, there is a significant increase in Prevotella copri, Eubacterium dolichum, Veillonella dispar, and Haemophilus parainfluenzae, along with a significant decrease in Anaerostipes [94]. Notably, Prevotella copri may be associated with susceptibility to inflammatory diseases by promoting the activation of Th17 cells and driving cytokines IL-6 and IL-23 [97]. Regarding CRC, the colonic microbiota has been extensively studied. Fusobacterium nucleatum in humans and mouse models produces the FadA adhesin binding to E-cadherin, which can activate the β-catenin pathway and promote the expression of transcription factors and oncogenes, thereby facilitating the growth of CRC cells [98,99,100]. Additionally, enterotoxin produced by B. fragilis induces the activation of activating transcription factor 3 to drive a Th17-type cellular immune response, which promotes inflammatory responses in the colonic mucosa in CRC by IL-23-driven production of IL-17A [101]. Therefore, F. nucleatum and B. fragilis are being explored as potential biomarkers for the diagnosis and treatment of CRC. Moreover, butyrate, a metabolite produced by the colonic microbiota, can induce G1 phase arrest or extensive apoptosis in CRC cells, which is an adjunct to conventional chemotherapy for CRC [102]. These findings suggest that further study of specific colonic microbiota will facilitate the development of new strategies for the identification, diagnosis, or treatment of IBD, IBS, and CRC.
Faecalibacterium, a key genus within the colonic microbiota, is associated with colitis—a complication of various diseases. For example, in the colonic MAM of Parkinson's disease (PD) patients, the relative abundance of Faecalibacterium (anti-inflammatory, butyrate-producing microbe) is significantly reduced, while Ralstonia (pro-inflammatory Proteobacteria) is increased. This imbalance may contribute to intestinal inflammation, α-synuclein misfolding, and subsequent neuroinflammation in PD [103, 104]. These observations are consistent with studies indicating that PD patients often exhibit colonic inflammation, which affects neuroinflammation via the brain-gut axis [105, 106]. In another study, Lou Ming [107] found in sleep-deprived mouse models, there was a decrease in the diversity and abundance of colonic microbiota, especially probiotics such as Akkermansia, Bacteroides, and Faecalibacterium, alongside a significant increase in pathogens like Aeromonas. This microbial shift may contribute to colonic mucosal damage. Interestingly, melatonin treatment upregulated Faecalibacterium abundance and its metabolite butyrate, thereby preventing colitis induced by acute sleep deprivation in mice [108, 109]. In addition, Anthony [110] demonstrated that changes in the anti-inflammatory status mediated by Faecalibacterium in the colonic microbiota may be a key factor in poor oral health. Individuals with more tooth loss exhibited a lower relative abundance of Faecalibacterium and a higher relative abundance of Proteobacteria in the colonic MAM [110]. These findings highlight the critical role of Faecalibacterium in the colonic microbiota in the development of various colitis-associated diseases, including PD, sleep deprivation-related conditions, and oral diseases.
Disturbance in the colonic microbiota can also lead to the formation of calcium oxalate kidney stones and hepatic encephalopathy. For example, Oxalobacter formigenes, a member of the colonic microbiota, degrades oxalate to obtain energy, thereby reducing intestinal oxalate secretion and urinary oxalate excretion. This makes it a potential as probiotics for the treatment of hyperoxaluria and calcium oxalate kidney stones [111,112,113]. Additionally, a decrease in diversity of the colonic MAM and significantly lower numbers of Bacteroides contribute to the development of mild hepatic encephalopathy in cirrhotic patients through gut-liver-brain axis [114, 115]. These findings confirm that the colonic microbiota can modulate the progression of various diseases through multiple gut-organ axes.
In summary, each intestinal segment possesses unique physiological functions and distinct microbiota compositions that collectively contribute to overall gut homeostasis and host health. Understanding the specific interactions between microbiota and their respective intestinal environments is essential for elucidating their roles in various physiological and pathological processes. This knowledge lays the foundation for developing targeted microbiome-based diagnostic and therapeutic strategies for a wide range of diseases.
Rectal microbiota and diseaseAlterations in the rectal microbiota have been implicated in various gynecological and systemic diseases. For example, Azpiroz [116] observed a significant reduction in rectal microbiota diversity in infertile women. Specifically, there was an increased Firmicutes/Bacteroidetes ratio and a decreased relative abundance of Prevotella, which is essential for producing short-chain fatty acids (SCFAs) that help construct the intestinal mucosal barrier. Concurrently, an increase in Bacteroides, which can prevent intestinal mucus synthesis by producing metabolites, was noted. These changes collectively weaken the rectal mucosal barrier in infertile women [117]. Additionally, infertile women experiencing repeated implantation failure exhibited an elevated presence of Erysipelotrichaceae in their rectal microbiota. This bacterial family induces TNF-α-driven inflammation and insulin-resistant obesity, ultimately contributing to implantation failure [117].
The anatomical proximity between the female urogenital tract and the anorectal region facilitates microbial translocation, potentially linking rectal microbiota alterations to female infertility [118]. Supporting this, the presence of Prevotella in both rectum and genital tract has been associated with an increased risk of genital inflammation and HIV infection [119]. These findings suggest that the interaction between rectal microbiota and urogenital microbiota may become a focal point for future research in gynecological diseases.
Beyond gynecological conditions, rectal microbiota is connected to other health issues. In patients with community-acquired pneumonia (CAP) hospitalized for one month, rectal swab analysis revealed that changes in the rectal microbiota influenced cytokine levels and degranulated products. Specifically, alterations in the abundance of butyrate-producing bacteria were associated with changes in IL-27 and IFN-ɣ expression, potentially leading to recurrent infections and rehospitalization post-CAP recovery [120]. Furthermore, David [121] utilized rectal swabs to examine the rectal microbiota of patients with alcohol-associated cirrhosis, finding a significantly lower abundance of Enterobacteriaceae, particularly Escherichia coli, compared to non-alcohol-associated cirrhosis. This suggests that Enterobacteriaceae characteristics could differentiate cirrhosis based on etiologies.
Yu-Fei [122] conducted rectal biopsies in patients with subclinical ulcerative colitis (UC) and identified an increase in Bacteroidetes, Blautia, Ruminoccocaceae, and Lachnospiraceae, alongside a decrease in Proteobacteria, Bacteroides, Prevotella, and Faecalibacterium in their rectal MAM. These microbial patterns may serve as biomarkers to distinguish subclinical UC from healthy individuals. Additionally, Shanshan [123] discovered that in patients with acute pancreatitis (AP), the rectal microbiota exhibited elev
Comments (0)