Sambucus williamsii Hance (SWH) belongs to Caprifoliaceae family and is widely distributed in China, Korea and Japan [13]. It is in the same genus as elderberry (also known as Sambucus nigra) which is popular in Europe to be used in products of syrup, gummies, soft drink, liqueur and eye gel. S. williamsii was first recorded in Tang Materia Medica (A.D. 659) for healing fractures and alleviating pain. Our previous studies have demonstrated that the SWH extract and its lignan-rich fraction increased BMD, improved trabecular bone micro-architecture, and promoted cortical bone strength in ovariectomized mouse and rat models [14,15,16,17]. Fifty-six lignans were isolated and purified in the extract of SWH and were identified to be the bioactive components in SWH through the determination of the osteoblastic proliferation in vitro [18, 19].
A Question Arises from Pharmacokinetics Study
In order to identify the major bioavailable lignans that are associated with the bone protective effects of SWH, a pharmacokinetic study using rat model was carried out. The rats were administrated with an effective dose or a 15-times effective dose of the SWH fraction, followed by collection of the blood samples at 0, 5, 15, 30, 60, 120, 180, 240, 360, and 480 min. However, neither the prototype nor metabolites were detected by highly sensitive method using an ultra-high performance liquid chromatography with quadrupole time-of-flight mass spectrometry (UPLC-QTOF/MS). To accumulate sufficient exogenous components in blood, samples were again collected and analyzed from rats gavaged daily with effective dose or 15-times effective dose of SWH fraction for 3 days. However, no detectable prototype nor metabolites of lignans could be detected in serum collected from these rats treated with SWH fraction for 3 days. The in-house results indicated that lignan-rich fraction may be composed of too many lignans with too low concentration of each type of lignan to be detected in blood. However, the lack of detectable lignans in the circulation might also suggest that mechanisms that do not require gastrointestinal uptake of lignans might be involved. This unsuccessful pharmacokinetic study prompted us to explore other mechanisms that might account for the bone protective effects of lignans.
Gut Microbiome Interaction with Lignan
Based on the fact that the bone protective effects of SWH was demonstrated in response to its oral administration, we hypothesized that lignans from SWH might exert its bone protective effect via the gut microbiome, including gut microbiota, their metabolites and their living environment [20]. An animal study using the lignan-rich fraction (CA) from SWH was performed to test the hypothesis [21]. Female SD rats (4-month old) were sham operated or ovariectomized, and treated with vehicle, PTH (1.8 µg/kg, n = 8, intramuscularly injected), CA (140 mg/kg, n = 10) for 10 weeks. The results exhibited that the lignan-rich fraction from SWH at 140 mg/kg prevented the reduction in BMD at distal femur and proximal tibia and improved bone microarchitecture in OVX rats, and these effects were similar to those of PTH on the trabecular bone.
The 16S rRNA sequencing of fecal samples showed that the microbial diversities of all groups increased significantly upon 10 weeks of treatment when compared to OVX group at 0 week, while no significant differences were found among the Sham, OVX and CA groups at week 10. The Principal Coordinate analysis (PCoA) and Partial least squares discriminant analysis (PLS-DA) analysis showed that the operational taxonomic units (OTUs) at week 10 were well separated from that at week 0, suggesting the microbial composition had altered over time. At week 10, the bacteria in the CA treatment group induced a significant separation from that of the Sham and OVX groups, indicating that CA treatment had significant influence on intestinal bacterial composition.
The difference analysis between groups and the LEfSe (linear discriminant analysis Effect Size) analysis showed that the abundance of Antinobacteria phylum extremely enriched in OVX rat fecal samples in response to CA treatment, and the taxa contributed to the abundance included: class, Antinobacteria; order, Coriobacteriales; family, Coriobacteriacene; genus, Adlercreutzia, norank_f_Coriobacteriaceae, Parvibacter, Enterorhabdus, Collinsella. The five genera belonged to Antinobacteria phylum were significantly reversed in the OVX rats treated with CA when compared with the OVX groups. These altered genera are related to the production of short chain fatty acid, modification of host bile acids and cholesterol levels, low dietary fiber intake and circulating insulin, cholesterol absorption, synthesis, and excretion [21]. Further spearman’s correlation analysis revealed that BMD was directly correlated with several genera, such as Lachnospiraceae_NK4A136_group and [Eubacterium] coprostanoligenes group, which are related to tryptophan metabolism. The results indicated that CA treatment might indirectly affect bone metabolism via modifying the metabolism of tryptophan in the gut.
Serotonin Involved in the Actions of Lignans on Bone
A metabolomic study was performed to identify if tryptophan metabolism was involved in the actions of lignans on bone. The resulted showed that tryptophan level in OVX rats was significantly restored upon treatment with the lignan-rich fraction of SWH for 12 weeks [17]. Tryptophan has two major catabolic pathways, the kynurenine (Kyn) pathway and serotonin (5-HT) pathway. The serum kynurenine level and serotonin level upon treatment with CA were quantified using an ultra-performance liquid chromatography tandem triple quadrupole mass spectrometry (UPLC-TQD-MS) method. The results showed that CA treatment significantly reduced serum 5-HT level, but not Kyn level, in OVX rats. The linear regression analysis showed that BMD was negatively correlated to serum 5-HT in rats [21].
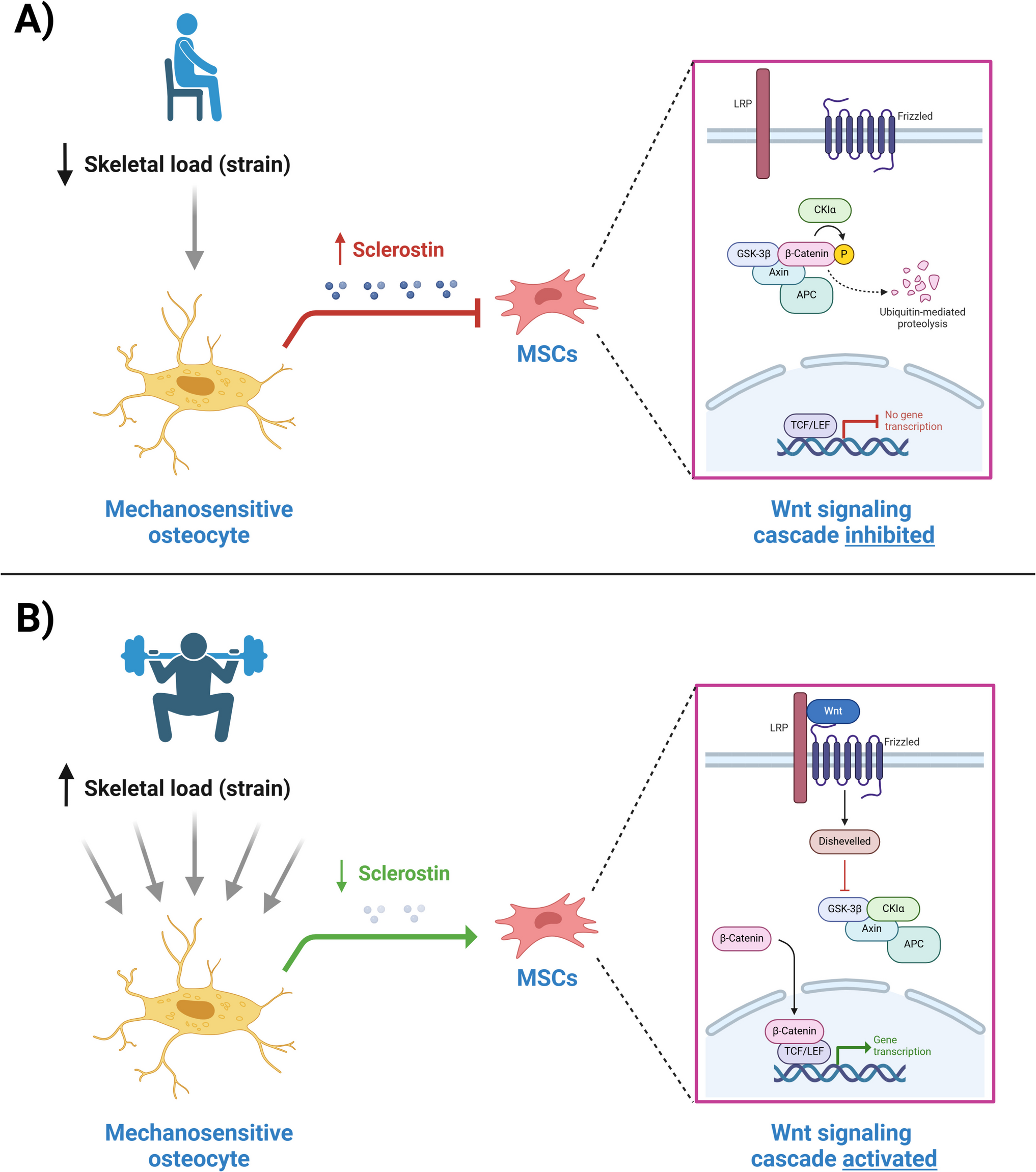
Serotonin is best known as a neurotransmitter, and more than 90% of the body’s serotonin is synthesized by enterochromaffin cells in intestine, while the others is synthesized by clusters of neurons in brain. As serotonin cannot pass through the blood–brain barrier, all serotonin in the circulatory system comes from intestine. Tryptophan hydroxylase (TPH) is the rate-limiting enzyme of 5-HT biosynthesis, and only the subtype of TPH-1 is located in the intestine, while the subtypes of TPH-1 and TPH-2 are distributed in the brain [22]. Our studies showed that CA significantly suppressed the protein expression of TPH-1 in the colon, while did not alter the protein expressions of TPH-1 and TPH-2 in cerebral cortex in OVX rats.
For the serotonin signaling pathway in osteoblast, the gut-derived 5-HT is delivered to osteoblasts by platelets, and its binding to the membrane 5HT1b receptor will then trigger the FOXO1/CRBE and FOXO1/ATF4 signaling cascade in osteoblastic cells. When high level of 5-HT acts on osteoblasts, the balance between FOXO1/CRBE and FOXO1/ATF4 will be broken and lead to the decrease in bone formation [23]. In our study, the mRNA and protein expressions of CREB in femur were not altered by OVX nor CA treatment. In contrast mRNA and protein expressions of ATF4 and FOXO1 in femur were significantly upregulated and downregulated in OVX rats, respectively; and these changes were significantly reversed in OVX rats in response to treatment with CA [21].
Taken together, our study clearly demonstrated that the lignan-rich fraction derived from Sambucus williamsii Hance exerted bone protective effects via altering circulating serotonin and gut microbiota.
Exploring a Novel Class of TPH-1 Inhibitor
Based on our discovery of the involvement of TPH-1 and serotonin in mediating the actions of lignan-rich fraction of SWH, subsequent study to identify TPH-1 inhibitors from SWH was performed [24]. The binding affinities of lignans isolated and purified from SWH were determined by molecular docking and surface plasmon resonance (SPR), the protein activity and expression of TPH-1 was tested in vitro, and serum serotonin level and BMD in mice upon treatment with purified lignan compound were measured. Our results showed that lignans derived from SWH exhibited high binding affinities to the TPH-1 protein, inhibited the activity and expression of TPH-1 in RBL2H3 cells, the Tph-1 gene high-expressing cells, and suppressed serum serotonin levels as well as significantly improved BMD in OVX mice. A series of novel class TPH-1 inhibitors, the lignans derived from SWH, were explored. The results provided the direct evidence that lignans could act on TPH-1 and influence serotonin synthesis and its circulating levels, thereby indirectly modulate bone metabolism in OVX animal model.




Comments (0)