
Remember me



Patients with type 2 diabetes suffer from increased susceptibility to infections, partly a direct consequence of hyperglycemia.1 2 Little is known about the effects of hyperglycemia on immune cell functions. In particular, peripheral blood neutrophils enriched from these patients exhibit a suboptimal response to activation that includes reduced pathogen killing, enhanced oxidative stress, and decreased apoptosis that may contribute to tissue injuries.3–6
Neutrophils are the first line of cellular defense against pathogens. These cells have a short lifespan and are unstable ex vivo, complicating their study post-enrichment. During their development, neutrophil progenitors reside in a hypoxic niche in the bone marrow and depend on glycolysis for energy production.7 On commitment to the neutrophil lineage, the mitochondria metabolism is activated and fueled by fatty acids oxidation.8 Circulating mature neutrophils contain few mitochondria and are mostly glycolytic, although these cells were recently evidenced as metabolically plastic.9–13 For example, while phagocytosis relies on glycolysis, the respiratory burst involves fast metabolic reorganization of glucose fluxes towards the pentose cycle. This enables high NADPH yields fueling NADPH oxidase (NOX) activity that boosts the oxygen consumption rate (OCR).14 15 It remains unclear how increased glucose availability in type 2 diabetes could impact neutrophil metabolism, differentiation, and functions.
Here we show that assessments of metabolic and functional features of donor neutrophils must be performed within a 3 hours window post-enrichment before significant alterations of metabolism and activation levels. As this short time frame is inadequate to evaluate multiple neutrophil functions and metabolic features, we present here an in vitro model of neutrophil-like cell differentiation in standard and higher glucose (HG) culture conditions to study the impact of glucose concentration on metabolism and functions of developing and mature neutrophils.
Research design and methodsDonor neutrophils enrichmentTen mL peripheral blood was collected into K2EDTA tubes (BD Biosciences) from healthy volunteers on signing informed consent. Anticoagulated blood was then diluted 1:1 with room temperature phosphate-buffered saline (PBS) (Gibco), layered on Ficoll-Paque PLUS (Cytiva), and centrifuged at 450 g for 30 min at 18°C. Donor neutrophil fractions were collected, and erythrocytes were lysed using 150 mM NH4Cl buffer. Enriched donor neutrophils were washed twice with PBS at 300 g for 5 min and resuspended in Roswell Park Memorial Institute Medium (RPMI) (Gibco) supplemented with 10% Fetal Bovine Serum (FBS) (Sigma-Aldrich) and 1× Antibiotic-Antimycotic (Gibco).
Cell cultureDonor neutrophils were incubated in RPMI/10% FBS at a density of 5×105 cells/mL over 3 hours at 4°C or 37°C, measurements were performed at 0, 1.5, and 3 hours. HL-60 cells (American Type Culture Collection (ATCC), CCL-240) were maintained in IMDM (Gibco) supplemented with 20% FBS and 1× Antibiotic-Antimycotic, as recommended by ATCC.16 All experiments were performed using passages ranging from 10 to 18. For differentiation towards neutrophil-like cells, HL-60 were seeded at 4×105 cells/mL in a medium supplemented with 1.3% dimethyl sulfoxide (DMSO) over 7 days, as previously described.17 Fresh medium was supplemented on day 4. For HG cultures, glucose (cat. G8270, Sigma-Aldrich) was supplemented to 50 mM to double the original glucose concentration.
Flow cytometryTitrated concentrations of the monoclonal antibodies against human CD11b (BD Biosciences, cat. 557918) and CD15 (BD Biosciences, cat. 563838) were used to investigate relative surface protein proportions on an FACSCelesta (BD Biosciences) fitted with 405 nm, 488 nm, and 640 nm lasers. Propidium iodide was added 15 min before acquisition as per the manufacturer’s instructions. For phagocytosis assays, opsonized Staphylococcus aureus BioParticles conjugated to Alexa Fluor 488 (AF488, Invitrogen) were prepared as per manufacturer’s instructions and incubated at 50:1 particles-to-cell ratio for 40 min at 37°C and 5% CO2. Cells were then washed with PBS and fluorescence was analyzed on the flow cytometer. For mitochondrial mass measurements, MitoTracker Green (Thermo Fisher, cat. M7514) was used at an optimized concentration of 200 nM. Fixable Viability Stain 510 (BD Biosciences, cat. 564406) was then incubated for 30 min. Cells were then washed with PBS and analyzed on the flow cytometer. For all flow cytometry data, at least 10,000 live singlets were analyzed per sample. Data were analyzed using FlowJo V.10 (BD Biosciences).
OCR measurementsOCR was measured in real time using a high-resolution respirometer (Oroboros O2K oxygraphy, Innsbruck, Austria). Briefly, 5×105 donor neutrophils/mL or 7.5×105 HL-60 or differentiated HL-60 (dHL-60)/mL were suspended in 2 mL of culture medium with a constant stirring at 500 rpm and 37°C. All OCR measurements started after a steady state was reached with at least 5 min routine respiration recording prior to adding relevant reagents as follows (online supplemental figure 1). 50 ng/mL phorbol 12-myristate 13-acetate (PMA) was added to reach the maximum OCR (OCRMax). The maximum acceleration of the OCR was calculated from the maximum slope between consecutive points of OCR tracings (figure 1A). To test NOX-specific OCR, 40 µM diphenyleneiodonium chloride (DPI, Sigma-Aldrich, cat. D2926) was added before or after PMA stimulation.
 Figure 1
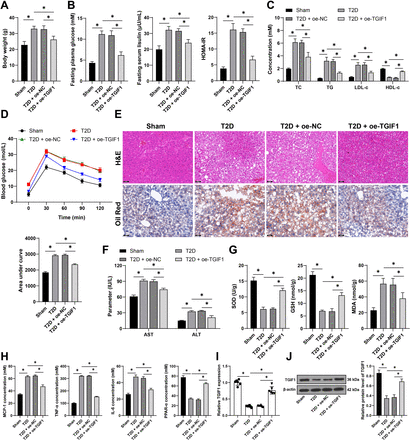
Figure 1 Ex vivo storage temperature and time elapsed post-enrichment regulate donor neutrophils OCRMax and CD11b surface expression. OCRMax is the maximum peak of the curve after PMA stimulation. (A) Donor neutrophils (n=5) were enriched and analyzed immediately (0 hours) or stored at 4°C (in blue) or 37°C (in red) and analyzed at 1.5 hours, and 3 hours post-enrichment for OCR post-PMA stimulation. (B) CD11b and CD15 relative cell surface expression were analyzed using flow cytometry on donor neutrophils directly post-enrichment (0 hours), or after 1.5 hours or 3 hours storage at 4°C (in blue) or 37°C (in red). Data is presented as biological replicates (A) or group mean (n=5)±SD for visual clarity (B). Two-way analysis of variance tests were used with Bonferroni post hoc to compare the impact of temperature at each time point and with Tukey post hoc to investigate changes across time within each group for each storage temperature. MFI, median fluorescence intensity; OCRMax, maximum oxygen consumption rate; PMA, phorbol 12-myristate 13-acetate.
For mitochondrial OCR, ATP synthase activity-dependent (ATP-linked) respiration was inhibited with 50 ng/mL oligomycin (Sigma-Aldrich, cat. 75351), followed by the addition of CCCP (carbonyl cyanyde 3-clorophenilhidrazone, Sigma-Aldrich, cat. C2759) to reach the maximum respiratory capacity. The optimal CCCP concentration was obtained by successive 0.5 µM additions of uncoupler (ranging 0.5–2 µM). Non-mitochondrial OCR was measured following the addition of 1 µM rotenone (Sigma-Aldrich, cat. 557368) and 2.5 µM antimycin A (Sigma-Aldrich, cat. A8674). Maximum electron transfer capacity (METC) is defined as the maximum respiratory capacity minus non-mitochondrial OCR. Spare capacity is defined as METC divided by routine respiration minus non-mitochondrial OCR (online supplemental figure 1).
To evaluate oxygen consumed by fatty acid oxidation, OCR was recorded over 40 min following the addition of 40 µM etomoxir (Sigma-Aldrich, cat. E1905). The final OCR was normalized with routine respiration. All data were normalized to live cell numbers manually counted immediately before OCR experiments using a Neubauer chamber with trypan blue exclusion.
Mitochondrial enzyme activityTo investigate relevant enzyme activities, cells were collected and washed with PBS before solubilization in lysis buffer (25 mM Tris-HCl, 150 mM NaCl, 0.5% NP-40). Homogenates were clarified by centrifugation at 7500 g. Citrate synthase activity was determined in total homogenates as previously reported.18 Carnitine palmitoyl transferase (CPT1) activity was measured as previously reported.19 Both activities were normalized to sample protein concentrations.
Morphological characteristicsMorphological characteristics of donor neutrophils were investigated using cytospin preparations. Briefly, 2×104 cells were spun onto poly-lysine coated slides at 1,100 rpm for 5 min using a Thermo Scientific Cytospin 4 centrifuge. Preparations were stained with Leishman’s stain and observed at 60× using a Ti Eclipse microscope (Nikon).
Statistical analysisData distributions were determined using Shapiro-Wilk tests. Two-way analysis of variance (ANOVA) tests were used to compare groups over time. One-way ANOVA tests were used to compare >2 groups. Relevant post hoc tests were applied to identify significant differences as described in figure legends. All statistical tests were performed using Prism V.9 software (GraphPad). P value<0.05 was considered statistically significant.
ResultsThe metabolism and activation status of donor neutrophils are both rapidly regulated ex vivoWe initially aimed to identify if time and storage conditions post venipuncture impacted neutrophil metabolism. We enriched donor neutrophils over a density gradient (mean CD15+CD11b+ purity of 89±6.08%, data not shown). The routine respiration of donor neutrophils immediately post-enrichment was null (online supplemental figure 2A–C). Adding PMA rapidly increased OCR to an OCRMax up to 83.61±19.63 pmol O2/(s×106 cells) (figure 1A). We confirmed OCR usage was directed to NOX by applying the NOX-specific inhibitor DPI pre-PMA and post-PMA stimulation (online supplemental figure 2A–C). We measured OCR immediately post-enrichment and after 1.5 hours and 3 hours at 4°C or 37°C, and obtained OCRMax. When donor neutrophils were stored at 4°C, there was a significant decrease in OCRMax 3 hours post-enrichment compared with freshly enriched donor neutrophils (p=0.0060, figure 1A). At 37°C, OCRMax also decreased between 0 hours and 3 hours (p=0.0287, figure 1A), and there was an additional significant reduction between 1.5 hours and 3 hours (p=0.0385). The time elapsed to reach OCRMax after PMA stimulation was unchanged (online supplemental figure 2D). Donor neutrophils stored at 37°C experienced a significant decrease in the OCRMax acceleration after PMA stimulation between 0 hours and 3 hours (p=0.0332, online supplemental figure 2E). Finally, we observed a reduction in nuclei and cytoplasm integrity after 3 hours at 4°C (online supplemental figure 2F). Overall, these findings suggest that donor neutrophils metabolism is regulated as early as 3 hours post venipuncture regardless of storage temperature. This is indicative of possible cell activation.
 Figure 2
Figure 2 Increasing glucose concentration during HL-60 neutrophil-like differentiation recapitulates some phenotypic features and functions of donor neutrophils. (A) Viability of HL-60 cells differentiated over 7 days with 1.3% dimethyl sulfoxide supplementation in 25 mM (yellow circles) or 50 mM glucose (orange squares). (B) Absolute fold expansion during HL-60 differentiation was calculated on days 2, 4, 5 and 7 using manual counting with trypan blue exclusion. (C) Proportion of CD11b+CD15+ cells in HL-60 and day 7-differentiated dHL-60 and HG-dHL-60 cultures. (D) Relative abundance of CD11b expressed as normalized MFI (ratio of stained over unstained controls, to account for distinct autofluorescence background) on the surface of HL-60 and day 7-differentiated dHL-60 and HG-dHL-60. (E) OCRMax post-PMA stimulation of HL-60 and day 7-differentiated dHL-60 and HG-dHL-60 cells. (F) Relative abundance of phagocytic cells in cultures of HL-60 and day 7-differentiated dHL-60 and HG-dHL-60, evaluated by flow cytometry as AF488+ cells post-AF488-Staphylococcus aureus BioParticles phagocytosis. Data presented as biological replicates (n=3–6 per group) and mean±SD. Two-way ANOVA with Tukey post hoc tests was used to investigate changes across time, and with Bonferroni post hoc tests to compare glucose concentrations in A–B. One-way ANOVA with Tukey post hoc tests was used to compare groups in C–F. *p<0.05, **p<0.01, ***p<0.001, ****p<0.0001. ANOVA, analysis of variance; dHL-60, differentiated HL-60; MFI, median fluorescence intensity; OCRMax, maximum oxygen consumption rate; PMA, phorbol 12-myristate 13-acetate.
To investigate cellular activation, we measured the stability of relevant proteins on the surface of donor neutrophils. We compared the relative expression of CD11b (integrin α-M) over 3 hours at 37°C or 4°C, as this informs on cellular activation.20 There was a significant increase in CD11b after 3 hours of incubation at 4°C, compared with freshly enriched donor neutrophils (p=0.0265, figure 1B). At 3 hours post-enrichment, CD11b was also significantly increased when cells had been stored at 4°C versus 37°C (p=0.0268). The expression of CD15, a characteristic marker for donor neutrophils, was not regulated by either time or incubation temperatures (figure 1B). Altogether, this is consistent with other reports of rapid ex vivo activation of donor neutrophils,21 22 urging the need for an in vitro model to study human neutrophils under specific conditions.
Higher glucose during HL-60 cell differentiation increases mature neutrophil-like cell yieldTo propose a surrogate for donor neutrophils and avoid ex vivo neutrophil regulations that could confound results, we used an established in vitro model of neutrophil-like cell differentiation using HL-60 cells stimulated with 1.3% DMSO (dHL-60). To evaluate the impact of hyperglycemia on neutrophil differentiation, we differentiated HL-60 cells in an HG media (HG-dHL-60). We first measured differentiation efficiency and cell viability in normal and HG. Cell viability was significantly decreased at day 4 in both glucose concentrations compared with HL-60 cells (p=0.0005 and 0.0493, for HG-dHL-60 and dHL-60, respectively, figure 2A). While decreased viability early in culture was evidenced in both differentiation protocols, HG-dHL-60 cultures yielded a significant increase in cell number at day 4 (p=0.0085, figure 2B) and dHL-60 showed a significant increase at day 7, versus starting concentrations (p=0.0075). At day 7, HG-dHL-60 yielded significantly more cells compared with dHL-60 (3.360 vs 2.508-fold increase, p=0.0435, figure 2B). These data demonstrate that HG correlates with increased cell yield.
Higher glucose during differentiation upregulates CD11b and increases phagocytosis without impacting ROSWe wondered if HG impacted neutrophil-like cells phenotypically. We evaluated relative CD11b and CD15 surface expression. Both dHL-60 and HG-dHL-60 exhibited the expected increase in CD11b+CD15+ cells at day 7 compared with HL-60 (p<0001, figure 2C). CD11b was significantly upregulated at day 7 for both culture conditions compared with HL-60 (p<0.0001 and p=0.0018, for HG-dHL-60 and dHL-60, respectively, figure 2D). Surprisingly, HG-dHL-60 had significantly increased CD11b compared with dHL-60 (p=0.0025, figure 2D). As CD11b increase is indicative of neutrophil activation, the measured increase in HG may be indicative of regulation of cellular functions.
To investigate the ability of neutrophil-like cells to produce reactive oxygen species (ROS), we measured OCRMax after PMA stimulation, as a surrogate for induced NOX activity (figure 2E). Unlike the rapid increase in OCR observed in donor neutrophils (online supplemental figure 2A), differentiated cells took an average of 20 min latency before a response could be registered post-PMA stimulation (online supplemental figure 3A). We confirmed OCR usage was directed to NOX in both glucose concentrations by applying DPI pre-PMA and post-PMA stimulation to differentiated cells (online supplemental figure 3). Only dHL-60 showed a significant increase in OCRMax compared with HL-60 cells (p=0.0241, figure 2E). Regarding phagocytosis, both differentiated cultures exhibited a significant increase in phagocytic cells (mean 26.98% and 50.77% for dHL-60 and HG-dHL-60, respectively) compared with HL-60 (mean 5.60%, p<0.0001 and p=0.006 for HG-dHL-60 and dHL-60, respectively, figure 2F). There were significantly more phagocytic cells in HG-dHL-60 cultures compared with dHL-60 (p=0.0003, figure 2F). Taken together, this suggests that increased glucose concentration during neutrophil differentiation increases phagocytosis without affecting ROS production in mature cells.
 Figure 3
Figure 3 Increasing glucose concentration during neutrophil-like HL-60 differentiation is associated with metabolic changes in the mitochondria. (A) Citrate synthase activity was measured in total protein lysates and normalized activity is presented in HL-60 and over a 7-day 1.3% dimethyl sulfoxide-induced neutrophil-like differentiation in 25 mM (dHL-60) or 50 mM (HG-dHL-60) glucose. (B) Relative mitochondrial mass was directly measured by flow cytometry using MitoTracker Green in HL-60 and on days 5 and 7 of neutrophilic differentiation in dHL-60 and HG-dHL-60. (C) Visual description of the specificity of electron transport chain inhibitors and of the membrane uncoupler CCCP (framed in red) that were used to measure metabolic changes in the electron transport chain, ATP synthase, and membrane proton flux (panel created using a modified template from BioRender). (D) Routine respiration was measured in the absence of an inhibitor or stimulus. (E) Non-mitochondrial OCR was measured during 5 min post-addition of rotenone and antimycin A in HL-60, dHL-60 and HG-dHL-60. (F) Maximum electron transport capacity (METC) was calculated as OCR post-CCCP addition minus non-mitochondrial respiration. Measurements in D–F were normalized to the number of live cells. Graphs showing three to five biological replicates per group, and group mean±SD. Two-way analysis of variance with Tukey post hoc tests was used to investigate changes across time, and with Bonferroni post hoc tests to compare glucose concentrations. *p<0.05, **p<0.01, ***p<0.001, ****p<0.0001. CCCP, carbonyl cyanyde 3-clorophenilhidrazone; dHL-60, differentiated HL-60; HG, higher glucose; MFI, median fluorescence intensity; OCR, oxygen consumption rate.
Higher glucose downregulates citrate synthase activity at days 5 and 7 of differentiation without impacting mitochondrial massHaving established that an HG concentration during neutrophil-like differentiation impacts mature cell functions, we wanted to investigate a possible link with the metabolism of differentiating cells. We measured mitochondrial mass during neutrophil-like differentiation using MitoTracker Green, which selectively accumulates on the mitochondrial matrix where it covalently binds to mitochondrial proteins reacting with free thiol groups from cysteine residues. MitoTracker Green median fluorescence intensity values decreased significantly between undifferentiated cultures and day 5 (p=0.0010 and p=0.0004 for dHL-60 and HG-dHL-60, respectively, figure 3B) and day 7 (p=0.0007 and p=0.0003 for dHL-60 and HG-dHL-60, respectively) of differentiation, but there was no change dependent on glucose concentration. Therefore, the mitochondrial mass decreased during neutrophilic differentiation, as previously reported,23 but this was irrespective of glucose concentrations.
We additionally measured citrate synthase activity which has been commonly used as a surrogate for mitochondrial mass.24–26 Citrate synthase is embedded in the mitochondrial matrix and is the first enzyme to participate in the tricarboxylic acid cycle. We evidenced significant reductions in citrate synthase activity in the HG environment, as early as day 2 (p=0.0037, figure 3A) and until day 7 (p=0.0003) of differentiation, compared with undifferentiated HL-60. On the other hand, there was no change in citrate synthase activity throughout differentiation in standard glucose (figure 3A). In mature neutrophil-like cells, citrate synthase activity was significantly reduced in the HG environment (p=0.0011 and 0.0034 for days 5 and 7 of differentiation, respectively, figure 3A). Overall, the mitochondrial mass decreased during neutrophil-like differentiation irrespective of glucose concentrations. On the other hand, citrate synthase activity was regulated independently of mitochondrial mass, and HG led to its downregulation in differentiating and mature neutrophil-like cells.
Mitochondrial oxygen consumption during neutrophil-like differentiation decreases irrespective of glucose concentrationAs citrate synthase activity was impaired in HG, we hypothesized a consequence in mitochondrial respiration during differentiation. We measured OCR while selectively impairing mitochondrial processes during neutrophilic differentiation in standard and HG (figure 3C). Routine respiration was significantly reduced from day 2 of differentiation irrespective of glucose concentration (p<0.0001, figure 3D), and this reduction remained stable until day 7 (p<0.0001). Non-mitochondrial OCR remained stable throughout the 7-day differentiation in both culture conditions (figure 3E). Consistent with routine respiration and mitochondrial mass decrease, METC also decreased on day 2 compared with undifferentiated cells (p=0.0002 and 0.0089 for dHL-60 and HG-dHL-60, respectively, figure 3F). METC is a parameter that informs on the maximum capacity of the mitochondria electron transport chain. HG showed a decreasing trend as day 5 and 7 had a significant decrease compared with day 2 (p=0.0066 and p=0.0056 for day 5 and 7, respectively, figure 3F) and day 4 (p=0222 and p=0.0191 for day 5 and 7, respectively). Additionally, we observed an increase in METC at day 4 in HG compared with dHL-60 (p=0.0429). Spare respiratory capacity, which estimates the unused but available capability to direct OCR to ATP production, did not show any significant difference compared with HL-60 either between glucose concentrations (online supplemental figure 4A). However, there was no change between conditions suggesting that cells could respond to stressors regardless of glucose concentration. ATP-linked respiration, which informs on OCR-mediated ATP production, was significantly decreased from day 2 onwards, irrespective of glucose concentration (p=0.0288 and 0.0340 for dHL-60 and HG-dHL-60, respectively, online supplemental figure 4B). Altogether, this suggests that differentiation causes mitochondrial activity impairment. However, glucose concentration may not impair this process further.
 Figure 4
Figure 4 Regulation of fatty acid oxidation during neutrophil-like HL-60 differentiation. (A) The contribution of fatty acid oxidation (FAO) to energy production during neutrophil-like cell differentiation was measured in HL-60 (day 0) and at days 2, 4, 5 and 7 of differentiation in dHL-60 and HG-dHL-60. Etomoxir (framed in red) inhibits CPT1 which is required to transport fatty acids inside the mitochondria for oxidation to occur. OCR inhibited by etomoxir was normalized with routine respiration and is represented as a ratio. (B) CPT1 activity was measured in total protein lysates. Data is presented as individual biological replicates (n=2–4 per group) and group mean±SD. Two-way analysis of variance tests were used with Bonferroni post hoc to compare the impact of glucose concentration at each time point, and Tukey post hoc was used to investigate changes across time for each glucose concentration. *p<0.05, ***p<0.0001. CoA, coenzyme A; CPT1, carnitine palmitoyl transferase; dHL-60, differentiated HL-60; OCR, oxygen consumption rate.
Higher glucose downregulates CPT1 activity during neutrophil-like cell differentiationWe further investigated mitochondrial processes by focusing on fatty acid oxidation, a process recently involved in healthy myeloid differentiation.8 As HG provides a substrate for non-mitochondrial-mediated ATP production, we asked whether HG may also dysregulate fatty acid metabolism during differentiation. We measured OCR in the presence of etomoxir, a fatty acid oxidation inhibitor. There was no change between undifferentiated HL-60 and differentiating cells. At day 4 of differentiation, dHL-60 had significantly increased fatty acid oxidation compared with HG-dHL-60 (p=0.0351, figure 4A). At day 7, however, the trend was reversed with increased fatty acid oxidation-mediated ATP production in HG-dHL-60 (p=0.0277). We then measured CPT1 activity, the rate-limiting step in fatty acid oxidation (figure 4B). There was no difference between HL-60 and differentiating cells. However, there was a significant decrease in CPT1 activity in HG-dHL-60 compared with dHL-60 starting day 4 and throughout day 7 (p=0.0139, p<0.0001, and p=0.0167, for days 4, 5, and 7, respectively, figure 4B). Overall, these data suggest that HG decreases CPT1 activity, leading to lower fatty acid oxidation abundance during neutrophilic differentiation.
ConclusionsWe describe here DMSO-differentiated HL-60 cells cultured in HG as a surrogate in vitro model to study hyperglycemic neutrophils and their differentiation metabolism. The model displays metabolic features similar to those reported from differentiating and mature primary neutrophils.17 23 27 28
HG caused a higher yield of neutrophil-like cells at day 7 of the differentiation. Hyperglycemia has previously been correlated with increased expression of receptors for advanced glycation end products (RAGE) in common myeloid progenitors in a mouse model, eventually promoting myeloid cell production.29 RAGE are stimulated by S100A8/A9 proteins secreted by neutrophils. This leads to colony factor secretion favoring granulocyte-macrophage progenitor proliferation, increasing neutrophil production. We suggest increased glucose may promote early mature neutrophils to enhance RAGE in late progenitors, fueling neutrophil progenitors’ proliferation.
In differentiated cells, CD11b was upregulated in HG, consistent with a report of hyperglycemic donor neutrophils.30 This phenomenon was attributed to a failure in actin polymerization after neutrophil activation in patients with type 2 diabetes, provoking a persistent CD11b expression combined with reduced extravasation.31 We also measured more phagocytic cells in HG cultures. Others have similarly described an increase in phagocytic pathway usage in hyperglycemic donor neutrophils using phosphoproteomics.32 Therefore, increasing glucose concentration in HL-60 cell differentiation at least partially correlated with the phenotype of neutrophils from patients with hyperglycemic type 2 diabetes.
In HG, citrate synthase activity was downregulated within the available mitochondria during differentiation. As mitochondrial mass was similar in both conditions, HG could downregulate citrate synthase activity within the available mitochondria. To the best of our knowledge, this is the first report of citrate synthase activity regulation independent of mitochondrial mass. Therefore, at least in this model, citrate synthase activity measurement may not be an appropriate surrogate for mitochondrial mass relative assessment, which has also been suggested in a mouse study.33 In addition, during differentiation energy is derived from fatty acid oxidation, facilitated by the CPT1 transport of fatty acids into the mitochondria.8 Here, we show that HG causes a downregulation of CPT1 throughout differentiation. HG promotes malonyl-coenzyme A (CoA) production, which has been shown to inhibit CPT1.34–36 We suggest this could be a mechanism for the observed CPT1 reduction in HG. The unexpected but consistent decrease in OCR post-etomoxir at day 7 in HG may be artifactual, as etomoxir also inhibits complex I of the electron transport chain and globally reduces OCR.37
Despite recapitulating various features of hyperglycemic human neutrophils, we were unable to directly compare the model with donor neutrophils in the study time frame. This said, we also demonstrated that donor neutrophils are rapidly regulated ex vivo which may confound results interpretation. We acknowledge the supraphysiological glucose concentration of 25 mM that is recommended as standard for the culture of HL-60 cells and used as the lower glucose concentration here.16 Another possible limitation of this model is the normoxic environment (5% CO2) in sharp contrast to the hypoxic niche where primary neutrophils are produced. Of note, neutrophils have been successfully produced from primary CD34+ hematopoietic stem cells in normoxia ex vivo.38 In the context of evaluating the impact of glucose on neutrophil differentiation and metabolism, we describe that HL-60 differentiated neutrophil-like cells may be used to unearth novel metabolic features of differentiation in HG, such as the regulation of citrate synthase independent of mitochondrial mass.
Comments (0)