
Remember me


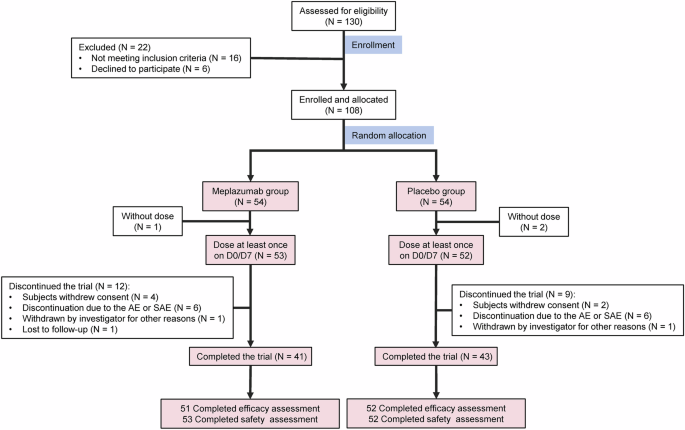



We chose to develop a mouse model with a gain-of-function mutation in PIK3CA that specifically targets endothelial cells. For this purpose, we utilized the transgenic mouse strain R26StopFLP110* that is designed to express a dominant active PIK3CA transgene upon Cre recombination. By crossing these mice with Tie2 Cre mice, we thus generated PIK3CATie2-CreER animals that carry a constitutively overactivated form of PIK3CA in capillaries and vein endothelial cells upon tamoxifen administration. To monitor Cre recombination, we further bred the PIK3CATie2-CreER mice with Gt(ROSA)26Sortm4(ACTB-tdTomato,-EGFP)Luo/J mice in which an ubiquitously expressed cell membrane-localized tdTomato fluorescent protein is replaced by GFP following Cre recombination.22 For control purposes, littermate mice carrying only the Tie2 Cre recombinase without the PIK3CA transgene were used and will be referred to hereafter as PIK3CAWT mice.
To address developmental concerns, we utilized four-week-old mice and administered a daily dose of 40 mg.kg-1 tamoxifen for five consecutive days to induce Cre recombination. After observing a delay of 2 to 3 weeks following Cre recombination, we noticed progressive and visible generalized subcutaneous capillary and venous malformations in PIK3CATie2-CreER mice (Fig. 1a), accompanied by excessive weight gain (Supplementary Fig. 1a). Both male and female mice exhibited these characteristics (Supplementary Fig. 1a). The malformations showed rapid growth and were associated with a significant reduction in survival compared to controls (Fig. 1b). To further investigate the extent of the malformations, we conducted whole-body T2-weighted magnetic resonance imaging (MRI) at 5 weeks after Cre induction. These studies revealed the presence of disseminated superficial capillaries and deep venous malformations (Fig. 1c and Supplementary Fig. 1b). Necropsy examinations demonstrated subcutaneous hemorrhage all linked to disseminated vascular malformations in the peri-laryngeal space, lower and/or upper limbs, and urogenital area (Supplementary Fig. 1c).
Fig. 1
A mouse model of PIK3CA-related capillary venous malformations. a Representative photography of PIK3CAWT and PIK3CATie2-CreER mice 4 weeks after Cre recombination. b Kaplan–Meier survival curves of PIK3CAWT and PIK3CATie2-CreER mice (n = 25 per group). c Coronal whole-body T2-weighted fat saturated magnetic resonance images (MRI) of PIK3CAWT and PIK3CATie2-CreER mice (n = 4–6 mice per group) 5 weeks after Cre recombination. Volumetric quantification of the vascular malformations. d Representative hematoxylin and eosin (H&E) staining of the skin of PIK3CAWT and PIK3CATie2-CreER mice and patient with venous malformation (VM). Large irregular and dilated vessels filled with red blood cells were visible in the skin of PIK3CATie2-CreER mice. Scale bar: 40 μm. e Representative coimmunofluorescence staining of Tie2 and GFP in the skin of PIK3CAWT and PIK3CATie2-CreER mice. Scale bar: 10 μm. f GFP is express in CD31+ cells as assessed by flow cytometry experiments of cells isolated from the skin of PIK3CAWT (n = 4 mice). g GFP and podoplanin staining using flow cytometry experiments of cells isolated from the skin of PIK3CAWT (n = 4 mice). h Representative immunofluorescence staining of P-AKTThr308 and P-S6RP in the skin of PIK3CAWT and PIK3CATie2-CreER mice. Scale bar: 10 μm. i Western blot and j quantification of P-AKTSer473 and P-S6RP in the skin of PIK3CAWT and PIK3CATie2-CreER mice (n = 5–8 mice per group). k Flow cytometry experiments showing the percentage of GFP + CD31+ cells isolated from the skin of PIK3CAWT and PIK3CATie2-CreER mice expressing P-AKTSer473 (n = 6–7 mice per group). l Representative immunofluorescence staining of KI67 in the skin of PIK3CAWT and PIK3CATie2-CreER mice. Scale bar: 10 μm and m quantification (n = 4 mice per group). n Flow cytometry experiments showing the percentage of GFP + CD31+ cells isolated from the skin of PIK3CAWT and PIK3CATie2-CreER mice expressing KI67 (n = 3–4 mice per group). o Flow cytometry experiments showing the percentage of Tomato+ cells isolated from the skin of PIK3CAWT and PIK3CATie2-CreER mice expressing KI67 (n = 3–4 mice per group). p Quantification of GFP positive cell surface isolated from PIK3CAWT and PIK3CATie2-CreER mice (n = 5 mice per group). q Complete blood count and D-Dimers measurement in PIK3CAWT and PIK3CATie2-CreER mice (n = 11 per group)
Histological analysis of the capillary and venous malformations revealed dilated and irregular vessels with disorganized tissue structure, endothelial hyperplasia, blood-filled lumens, hemosiderin interposition, and features of hemophagocytosis (Supplementary Fig. 1d). In some mice, local clotting was accompanied by pulmonary embolism (Supplementary Fig. 1e, f). In the malformations, we also noticed varying degrees of adipose tissue overgrowth, lymphatic compounds, and αSMA positive cells (Supplementary Fig. 1f). These abnormalities closely resemble the ones that are seen in patients with capillary venous malformations (Supplementary Fig. 1g).
We then verified Cre recombination through GFP and p110* expression. Coimmunostaining (Fig. 1E and Supplementary Fig. 2a), Western blot (Supplementary Fig. 2b) and flow cytometry experiments confirmed that the Cre recombinase was specifically expressed in CD31 positive cells (97.8 ± 0.31% mean ± SEM, n = 4 mice). Conversely 0.54 ± 0.1% of the cells were GFP + CD31− showing indeed the endothelial cell expression of the Cre recombinase (n = 4 mice) (Fig. 1f). Importantly, no Cre expression was detected in lymphatic endothelial cells using flow cytometry experiments (0.67 ± 0.2%, mean ± SEM, n = 4 mice) (Fig. 1g). Indeed, the presence of adipose tissue overgrowth and lymphatic anomalies suggests potential communication between endothelial cells and surrounding tissues or cell recruitment. In this regard, we noticed infiltration of immune cells within the lesions as indicated by the presence of CD3+ cells (Supplementary Fig. 2c) and F4/80-expressing cells (that is, macrophages). Macrophages were visible either within the lumen or in the close vicinity of dilated vessels (Supplementary Fig. 2d–f), with a significant proportion of CD163, a receptor involved in clearing and endocytosis of hemoglobin/haptoglobin complexes (Supplementary Fig. 2d–f). Flow cytometry experiments showed that the Cre recombinase was not expressed in CD45+ (0.21±0.03% mean ± SEM, n = 4 mice) or CD34+(0.09±0.01%, mean±SEM, n = 4 mice) bone marrow cells (Supplementary Fig. 2g, h).
Coimmunofluorescence experiments using GFP, COUP-TFII, CD31, Podoplanin, Ephrin B2 and Endomucin on 30 μm sections revealed severe disorganization in the 3D structure of the skin vessels (Supplementary Figs. 3 and 4).
Immunofluorescence (Fig. 1h), Western blot (Fig. 1i, j) and flow cytometry studies (Fig. 1k) revealed activation of the AKT and mTOR pathways in the endothelial cells of PIK3CATie2-CreER mice by contrast to PIK3CAWT. Mechanistically, PI3Kα is known to activate cell growth and proliferation. We investigated cell proliferation using KI67 staining (Fig. 1l, m, Supplementary Fig. 5a, b) and flow cytometry experiments. In control mice, we noted that 1.7 ± 0.6% (mean ± SEM, n = 4) of CD31 + GFP+ and 11.5 ± 0.4% (mean ± SEM, n = 4) of the Tomato+ cells co-expressed KI67 (Fig. 1n, o). Remarkably, in PIK3CATie2-CreER mice we observed an elevated proliferation rate in the CD31 + GFP+ population (21.7 ± 6.2% were positive for KI67) as well as in Tomato+ cells (26.6±4.8% were positive for KI67) (Fig. 1n, o). This finding indicates the proliferation of endothelial cells and supports again a potential crosstalk between mutated and surrounding wild-type cells. Using Amnis ImageStream® system, we also found that mutated endothelial cells isolated from PIK3CATie2-CreER mice were hypertrophic by contrast to controls (Fig. 1p and Supplementary Fig. 5c).
Finally, complete blood counts in PIK3CATie2-CreER mice revealed significant regenerative anemia, a low platelet count, and elevated D-Dimers levels, consistent with the possibility of disseminated intravascular coagulation, as well as slight hypereosinophilia. All of these abnormalities are also consistent with what is observed in patients with venous malformations (Fig. 1q).23,24
We thus have successfully established a mouse model that accurately replicates the phenotype of patients who are affected by somatic or inherited forms of capillary venous malformations. This model should offer the advantage of providing mechanistic insight into both the PIK3CA- and TEK-related types of vascular diseases.
We then investigated whether the PIK3CA gain-of-function mutation in capillaries and veins would lead to detectable changes in circulating metabolites that could potentially serve as biomarkers for disease progression. For this purpose, we collected plasma samples from both PIK3CAWT and PIK3CATie2-CreER mice after a 12-hour fast and subjected them to thorough metabolomic analysis. PIK3CATie2-CreER mice were found to exhibit activated anabolic pathways, resulting in the accumulation of various amino acids, including arginine, histidine, phenylalanine, thymidine, threonine, 5-adenosyl-homocysteine, valine, orotic acid, acetyl-lysine, methyl-lysine, lysine, and oxoadipate, along with the metabolic intermediates of these amino acids. Additionally, there was evidence of increased mitochondrial respiration in these mice, as indicated by the plasma accumulation of succinic acid and acetyl-carnitine, and activation of the urea cycle as indicated by higher circulating levels of arginine and urate compared to controls (Supplementary Fig. 6). As such, circulating metabolites could serve as potential biomarkers for monitoring disease progression in PIK3CA-capillary venous malformations.
In humans, a great number of PIK3CA variants have been identified and found to exert varying effects on the recruitment of the AKT pathway.25 To simulate a higher degree of AKT recruitment, we also created PIK3CATie2-HO mice that bear homozygous PIK3CA mutations in capillaries and venous endothelial cells. After Cre recombination, a more severe phenotype emerged, characterized by the rapid occurrence of disseminated and voluminous capillary venous malformations with reduced life expectancy (Supplementary Fig. 7a–c). Whole-body T2-weighted MRI conducted five weeks after induction further confirmed the presence of extensive disseminated venous malformations (Supplementary Fig. 7d). Histological examination revealed severe widespread clotting and pulmonary embolism (Supplementary Figs. 7e and 4f), phosphorylation assays of tissues, robust activation of the AKT pathway (Supplementary Fig. 7g, h) and complete blood count, profound regenerative anemia (Supplementary Fig. 7i).
PIK3CA signals partially through AKT1 and not AKT2 in venous endothelial cellsIn mammals, AKT is encoded by three distinct genes: AKT1, AKT2, and AKT3, with AKT1 being the main isoform in endothelial cells. To explore whether PIK3CA gain-of-function mutations primarily signal via AKT or other downstream targets, we conducted a series of experiments using genetically modified mice.
First, we interbred AKT1flox/flox with PIK3CATie2R-CreER mice to create a PIK3CATie2R-CreER-AKT1KO model (referred to as PIK3CAAKT1KO). Upon inducing Cre induction, we observed that AKT1 deletion in capillaries and veins led to a moderate extension of lifespan (median survival of 80 days) and only delayed the occurrence of vascular malformations (Fig. 2a and Supplementary Fig. 8a) as MRI performed at 6 weeks still revealed diffused venous malformations (Fig. 2b and Supplementary Fig. 8b). Histologically, the malformations were similar to the ones observed in PIK3CATie2R-CreER mice (Fig. 2c). AKT pathway recruitment exhibited no reduction (Fig. 2d, e), and cell proliferation rates remained unaffected by deletion of AKT1 (Fig. 2f, g). AKT1 deletion did not rescue complete blood count anomalies either (Supplementary Fig. 8c). These findings suggest that the AKT1 isoform only plays a partial role in the PIK3CA-driven phenotype of vascular malformations.
Fig. 2
PIK3CA-related capillary venous malformations in PIK3CATie2-CreER mice only partially signal through AKT proteins. a Kaplan–Meier survival curves of PIK3CAWT, PIK3CATie2-CreER, PIK3CAAKT1KO, PIK3CAAKT2KO and PIK3CAAKT1AKT2-KO mice (n = 20 per group). b Coronal whole-body T2 weighted magnetic resonance images (MRI) of PIK3CAWT, PIK3CATie2-CreER, PIK3CAAKT1KO, PIK3CAAKT2KO and PIK3CAAKT1AKT2-KO mice 6 weeks after Cre recombination. Volumetric quantification of the vascular malformations (n = 4–6 mice per group). c Representative hematoxylin and eosin (H&E) staining of the skin of PIK3CAWT, PIK3CATie2-CreER, PIK3CAAKT1KO, PIK3CAAKT2KO and PIK3CAAKT1AKT2-KO mice. Scale bar: 10 μm. d Representative P-AKTSer473 in the skin of PIK3CAWT, PIK3CATie2-CreER, PIK3CAAKT1KO, PIK3CAAKT2KO and PIK3CAAKT1AKT2-KO mice. Scale bar: 10μm.*: Vascular malformation. e Western blot and quantification of P-AKTSer473 and P-S6RP in the skin of PIK3CAWT, PIK3CATie2-CreER, PIK3CAAKT1KO, PIK3CAAKT2KO and PIK3CAAKT1AKT2-KO mice (n = 4–5 mice per group). f Representative immunofluorescence staining of KI67 in the skin of PIK3CAWT, PIK3CATie2-CreER, PIK3CAAKT1KO, PIK3CAAKT2KO and PIK3CAAKT1AKT2-KO mice. Scale bar: 10 μm. g Flow cytometry experiments showing the percentage of GFP + CD31+ cells isolated from the skin of the different mouse models expressing KI67 (n = 3–4 mice per group). (n = 3–7 mice per group). h Western blot and quantification of AKT1 and AKT2 in vessels of PIK3CAWT, PIK3CATie2-CreER and PIK3CAAKT1AKT2-KO mice (n = 3–4 mice per group)
Next, we investigated the role of the AKT2 isoform by interbreeding AKT2−/− with PIK3CATie2R-CreER mice to obtain PIK3CATie2R-CreER-AKT2KO model (referred to as PIK3CAAKT2KO). Following the induction of Cre, we observed that PIK3CAAKT2KO mice exhibited the same clinical, biological, radiological and histological manifestations as PIK3CATie2R-CreER mice (Fig. 2a–c and Supplementary Fig. 8a, c). AKT phosphorylation in tissues and proliferation rates were also modestly affected in PIK3CATie2R-CreER animals (Fig. 2d, e). Thus, AKT2 deletion did not impact the PIK3CA-vascular malformation phenotype.
Finally, we investigated the simultaneous deletion of both AKT1 and AKT2 isoforms by interbreeding PIK3CAAKT1KO with PIK3CAAKT2KO mice. The resulting PIK3CAAKT1AKT2-KO pups were born in the expected Mendelian ratio and did not display any particular phenotype during the first few weeks of life. At 4 weeks of age, we activated Cre recombinase, and over time, PIK3CAAKT1AKT2-KO mice began to develop capillary and venous malformations (Supplementary Fig. 8a). Whole-body MRI performed 6 weeks after Cre recombination revealed diffuse malformations (Fig. 2b and Supplementary Fig. 8b). Similar to PIK3CAAKT1KO mice, AKT1 and AKT2 deletion in capillaries and veins was associated with a moderate extension of lifespan (Fig. 2z). Histology and blood examination confirmed severe venous malformation with intravascular coagulation (Fig. 2c and Supplementary Fig. 8c). Once again, a modest reduction in AKT phosphorylation but not in the proliferation rate were observed (Fig. 2d–g). Western blot analysis showed that while AKT1 and AKT2 were successfully deleted in endothelial cells (Fig. 2h), AKT protein was still expressed, indicating possible compensation by AKT3. Indeed, we concluded that, genetic ablation of both isoforms AKT1 and 2 was associated with delayed occurrence of the malformations.
Targeted therapy for venous malformations in PIK3CA related venous malformationsWe proceeded to investigate whether targeted therapies.16 could improve the outcome of PIK3CATie2R-CreER mice. For this purpose, we compared the effects of three pharmacological inhibitors: rapamycin, miransertib, and alpelisib, targeting mTORC1,26 pan AKT, and PI3Kα, respectively. Four weeks after tamoxifen administration, mice were randomly assigned to one of the treatment groups.
Rapamycin, which is used off-label to treat various forms of venous malformations in patients, did not show any impact on the volume and histological aspects of venous malformations, on intravascular coagulation, or on overall survival when administered daily (Fig. 3a–d, Supplementary Fig. 9a, b). Rapamycin led to a reduction in S6RP phosphorylation in tissues, but AKT phosphorylation (Fig. 3e, f) and proliferation rate (Fig. 3g, h) remained very high.
Fig. 3
Targeted therapies for capillary venous malformations in PIK3CATie2-CreER mice. a Kaplan–Meier survival curves of PIK3CAWT and PIK3CATie2-CreER mice treated with either vehicle, rapamycin, miransertib or alpelisib (n = 12 per group). b Coronal whole-body T2-weighted magnetic resonance images (MRI) of PIK3CAWT and PIK3CATie2-CreER mice 6 weeks after Cre recombination treated with either vehicle, rapamycin, miransertib or alpelisib. Volumetric quantification of the vascular malformations (n = 3-4 mice per group). c Complete blood count in PIK3CAWT and PIK3CATie2-CreER mice treated with either vehicle, rapamycin, miransertib or alpelisib (n = 3–5 mice per group). d Representative hematoxylin and eosin (H&E) staining of the skin of PIK3CAWT and PIK3CATie2-CreER mice treated with either vehicle, rapamycin, miransertib or alpelisib. Scale bar: 10 μm. e Representative P-AKTThr308 and P-S6RP immunostaining in the skin of PIK3CAWTand PIK3CATie2-CreER mice treated with either vehicle, rapamycin, miransertib or alpelisib. Scale bar: 10 μm. f Western blot and quantification of P-AKTSer473 and P-S6RP of the skin of PIK3CAWTand PIK3CATie2-CreER mice treated with either vehicle, rapamycin, miransertib or alpelisib (n = 3–4 per group). g Representative immunofluorescence staining of KI67 in the skin of PIK3CAWTand PIK3CATie2-CreER mice treated with either vehicle, rapamycin, miransertib or alpelisib. Scale bar: 10 μm. h Proliferation index quantification (n = 3–4 mice per group)
Miransertib, which had shown encouraging preclinical results,16 resulted in a slight reduction in venous malformation volumes (Fig. 3b, Supplementary Figs. 9a, b) and modest lifespan extension (Fig. 3a). However, it did not significantly impact platelet level, anemia or the histological abnormalities (Fig. 3c, d). Western blot and immunofluorescence studies did reveal a reduction in AKT and S6RP phosphorylation (Fig. 3e, f) with a modest impact on proliferation (Fig. 3g, h). These findings suggest again that PIK3CA-capillary venous malformations are not solely dependent on AKT for their development
In contrast to these 2 previous drugs, alpelisib showed significant efficacy. It led to prolonged survival (Fig. 3a), a notable reduction in venous malformations volume as confirmed by MRI (Fig. 3b, Supplementary Fig. 9a, b) and histological analyses (Fig. 3d) and to the reversal of intravascular coagulation biologically (Fig. 3c). In the affected tissues, the AKT pathway and cell proliferation were completely inhibited (Fig. 3e–h). These results demonstrate that alpelisib outperformed rapamycin and miransertib in treating PIK3CATie2R-CreER mice, showing promising potential as a therapeutic option for the management of vascular malformations in particular.
Alpelisib prevents venous malformations in PIK3CA mouse modelWe next aimed to assess whether alpelisib could be effective in both preventing and improving vascular malformations in PIK3CATie2-CreER mice. To conduct a preventive study, we began to administer alpelisib vs. a vehicle 48 h after Cre induction and maintained it for 12 weeks. Alpelisib-treated PIK3CATie2-CreER mice displayed a visibly normal appearance and weight gain profile with extended lifespan compared to vehicle-treated PIK3CATie2-CreER mice (Fig. 4a, Supplementary Fig. 10a, b). MRI conducted after 6 weeks of alpelisib demonstrated the absence of vascular malformations (Fig. 4b, Supplementary Fig. 10c, d) and similar conclusions could be drawn based on necropsy examination (Fig. 4c). Alpelisib also effectively inhibited AKT and mTOR (Fig. 4d, e and Supplementary Fig. 10e), improved vascular anomalies (Supplementary Fig. 11a) and prevented blood cell count abnormalities from occurring (Fig. 4f).
Fig. 4
Alpelisib improves and prevent capillary venous malformations in PIK3CATie2-CreER mice. a Kaplan–Meier survival curves of PIK3CAWT and PIK3CATie2-CreER mice treated with either vehicle or preventive alpelisib (n = 12 per group). b Coronal whole-body T2-weighted magnetic resonance images (MRI) of PIK3CAWT and PIK3CATie2-CreER mice 6 weeks after Cre recombination treated with either vehicle, preventive or therapeutic alpelisib (n = 4–6 mice per group). Volumetric quantification of the vascular malformations. c Representative hematoxylin and eosin (H&E) staining of the skin of PIK3CAWT and PIK3CATie2-CreER mice treated with either vehicle, preventive or therapeutic alpelisib. Scale bar: 10 μm. d Representative P-AKTThr308 and P-S6RP immunostaining in the skin of PIK3CAWTand PIK3CATie2-CreER mice treated with either vehicle, preventive or therapeutic alpelisib. Scale bar: 10 μm. e Western blot and quantification of P-AKTSer473 and P-S6RP in skin of PIK3CAWTand PIK3CATie2-CreER mice treated with either vehicle, preventive or therapeutic alpelisib (n = 6–7 mice per group). f Complete blood count of PIK3CAWTand PIK3CATie2-CreER mice treated with either vehicle, preventive or therapeutic alpelisib (n = 5–10 mice). g Representative photography of PIK3CATie2-CreER mice before and two weeks after alpelisib initiation. h Kaplan–Meier survival curves of PIK3CAWT and PIK3CATie2-CreER mice treated with either vehicle or therapeutic alpelisib (n = 12 per group)
For the therapeutic study, alpelisib was administered to PIK3CATie2-CreER mice 4 weeks after Cre induction when vascular malformations were already visible. This treatment led to a rapid decrease in body weight in alpelisib-treated PIK3CATie2-CreER mice (Supplementary Fig. 10a), clinical improvement of the vascular malformations (Fig. 4g and Supplementary Fig. 10b), and extended lifespan (Fig. 4h). MRI performed 6 weeks after the start of alpelisib treatment showed a significant decrease in the volume of vascular malformations compared to vehicle-treated mice (Fig. 4b, Supplementary Fig. 10c, d). At the time of sacrifice, alpelisib-treated PIK3CATie2-CreER mice exhibited small vascular dilation (Fig. 4c), and AKT and S6RP phosphorylation were reduced (Fig. 4d, e and Supplementary Fig. 10e). Similar to the preventive study, blood anomalies and vessel malformations were corrected (Fig. 4f and Supplementary Fig. 11a). Furthermore, we investigated the impact of alpelisib on the plasma metabolites identified as dysregulated in PIK3CATie2-CreER mice. These metabolites completely or partially normalized under alpelisib in both the preventive and therapeutic conditions (Supplementary Figs. 12 and 13), indicating that they hold potential as early biomarkers of disease progression or regression in the mouse model. Alpelisib had no particular impact on the control mice. Finally, to explore whether vascular malformations could recur following the interruption of alpelisib, we induced Cre recombination in 4 male mice, treated them with alpelisib at 8 weeks post tamoxifen induction when they were severely affected for a period of 16 weeks, and then stopped the drug. In the following weeks, the mice restarted to gain body weight with the development of diffuse vascular malformations (Supplementary Fig. 11b). Indeed, alpelisib halted disease development but did not correct the genetic anomaly.
Alpelisib improves capillary venous malformations in patients with PIK3CA or TEK variantsAfter obtaining encouraging results, we received authorization from the French regulatory agency (ANSM) and The Comité de convenance du CHU de Québec to treat a total of 25 patients, both pediatric and adult, who had venous malformations associated with either PIK3CA mutation (n = 16) or Tie2R (TEK) mutation (n = 9), all demonstrating AKT pathway recruitment within the capillary venous malformations (Fig. 5a–c). However, we observed no difference in KI67 staining between controls and patients (Supplementary Fig. 14). The clinical and demographical characteristics of these patients are summarized in Tables 1 and 2, but briefly there were 10 women, 7 pediatric patients and the mean age was 29.12 years old (SD ± 15.49). All patients were previously heavily treated with multiple surgical, radiological procedures and treatment based on rapamycin for a minimum period of time of 6 months (Tables 1 and 2). Patients were selected because of rapamycin inefficiency (disease progression or uncontrolled symptoms or serious adverse events). All patients underwent a Doppler ultra sound and an MRI before initiating treatment with alpelisib and then a second MRI approximately 6 months after starting the medication. The dosage of alpelisib administered ranged from 50 to 250 mg, adjusted according to age based on the EPIK P1 clinical trial.21 After the introduction of alpelisib, all patients experienced improvements in symptoms such as pain or bleedings. Furthermore, there was a noticeable discoloration and reduction in the volume of vascular malformations observed in the patients (Tables 1 and 2, Fig. 5d, e and Supplementary Fig. 15). Specifically, PIK3CA-related venous malformations showed a mean reduction of 33.4% (SD, ± 22.1) from the baseline at 6 months, while TEK-related venous malformations demonstrated a mean reduction of 27.8% (SD, ±18.9) (Tables 1 and 2, Fig. 5f and Supplementary Fig. 16). Finally, we explored the metabolomic changes in patients following alpelisib introduction. Notably, we identified changes in multiple metabolites that aligned with the ones observed in the mouse model such as aspartate, cystine, glycerol, guanosine, inosine or succinic acid (Supplementary Fig. 17). These metabolic compounds could potentially serve as the foundation for disease improvement biomarkers.
Fig. 5
Alpelisib improves patients with PIK3CA or TEK-related capillary venous malformations. a Representative Hematoxylin and eosin (H&E) staining and b Immunofluorescence of P-AKTThr308 and P-S6RP in skin biopsies performed in controls and in patients with PIK3CA-related capillary venous malformation. Scale bar: 10 μm. c Immunofluorescence quantification (n = 6 controls and 6 patients). AU Arbitrary units. d Representative photographs of the morphological changes observed in patients with PIK3CA-related capillary venous malformations receiving alpelisib for 6 months. e Representative photographs of the morphological changes observed in patients with TEK-related capillary venous malformations receiving alpelisib for 6 months. f Transversal (upper panel) and coronal (lower panel) T2-weighted fat saturated MRI sequence of patient 1 and 5 before and after alpelisib introduction. In red, segmentation in 2D (left panel) and 3D (right panel). g Percentage change of the volume of the preselected lesion in patients with PIK3CA-related capillary venous malformations (left panel) and TEK-related capillary venous malformations (right panel)
Table 1 Patient characteristics with PIK3CA-related venous malformationsTable 2 Patient characteristics with TEK-related venous malformations
Comments (0)