
Remember me








Oxidative stress is characterized by an imbalance between the production of reactive oxygen species (ROS) and the body’s ability to neutralize these harmful substances. It plays a pivotal role in the development of metabolic syndrome, including heart failure and insulin resistance. In heart failure, an excess production of ROS relative to antioxidant defense can lead to cardiac dysfunction, contributing to the pathophysiology of the condition [1]. Insulin resistance, a key component of metabolic syndrome, has also been linked to oxidative stress. Increased oxidative stress can lead to insulin resistance, dyslipidemia, β-cell dysfunction, and impaired glucose tolerance, ultimately leading to type 2 diabetes (T2D) [2]. Insulin resistance in the myocardium can generate damage through altered signal transduction, impaired regulation of substrate metabolism, and altered delivery of substrates to the myocardium. These alterations can contribute to cardiomyopathy and heart failure [3].
Natriuretic peptides are family of neurohormones that are genetically discrete and mainly synthesized by the heart and the brain [4]. N-terminal pro B-type natriuretic peptide (NT-proBNP) is one of the main types of natriuretic peptides released by cardiomyocytes as a counteraction to elevated stress on cardiac walls and vasoconstriction [5]. NT-proBNP is a prominent diagnostic marker of heart failure severity and cardiac dysfunction [6].
Studies have shown that assessing levels of NT-proBNP and oxidative stress markers may improve the detection of early stages of heart failure [7]. Indeed, reports have shown that lowered serum NT-proBNP was associated with high levels of carotenoids [8], superoxide dismutase, total antioxidant capacity, catalase [9], and ubiquinone [10]. The relationship between NT-proBNP and antioxidant metabolites is complex and multifaceted, with implications for the diagnosis, management, and prognosis of cardiovascular diseases [11]. Furthermore, the association between NT-pro-BNP and insulin resistance is inconsistent across the literature. While some recent studies suggest an inverse relationship between NT-proBNP and insulin resistance [12], others do not find this association [13]. Nevertheless, the interplay between NT-proBNP and antioxidant metabolites in the context of metabolic syndrome, such as insulin resistance, could be a potential area of focus for improving diagnostic and prognostic models in cardiovascular health. Metabolomics has a significant role, from understanding the metabolic pathways underlying heart failure and insulin resistance, to identifying potential biomarkers and therapeutic targets [14]. The comprehensive understanding of the interplay between oxidative stress, insulin resistance, and cardiovascular risk could potentially lead to the development of novel therapeutic strategies for metabolic disorders and cardiovascular diseases.
Our objective is to evaluate the association between circulating metabolic markers of oxidative stress and NT-proBNP levels in healthy individuals, then investigate this association after dichotomization of our cohort into insulin-resistant and insulin-sensitive individuals. This research aims to contribute to the development of improved diagnostic and prognostic models in the field of cardiovascular health.
Methods Data and study participantsThe study was approved by the Institutional Review Boards of the Qatar Biobank (QF-QBB-RES-ACC-00125). All participants provided informed consent. Data for 3000 participants was retrieved from Qatar Biobank (QBB), along with questionnaire related to medications, previous history of cardiovascular events, and laboratory measurements of clinical chemistry and endocrinology tests. Insulin resistance was determined by homeostatic model assessment of insulin resistance (HOMA-IR) calculated from fasting blood glucose (mmol/l) and fasting insulin mIU/ml measurements using the following equation:
HOMA-IR=Insulin×Glucose(mmol/l)/22.4
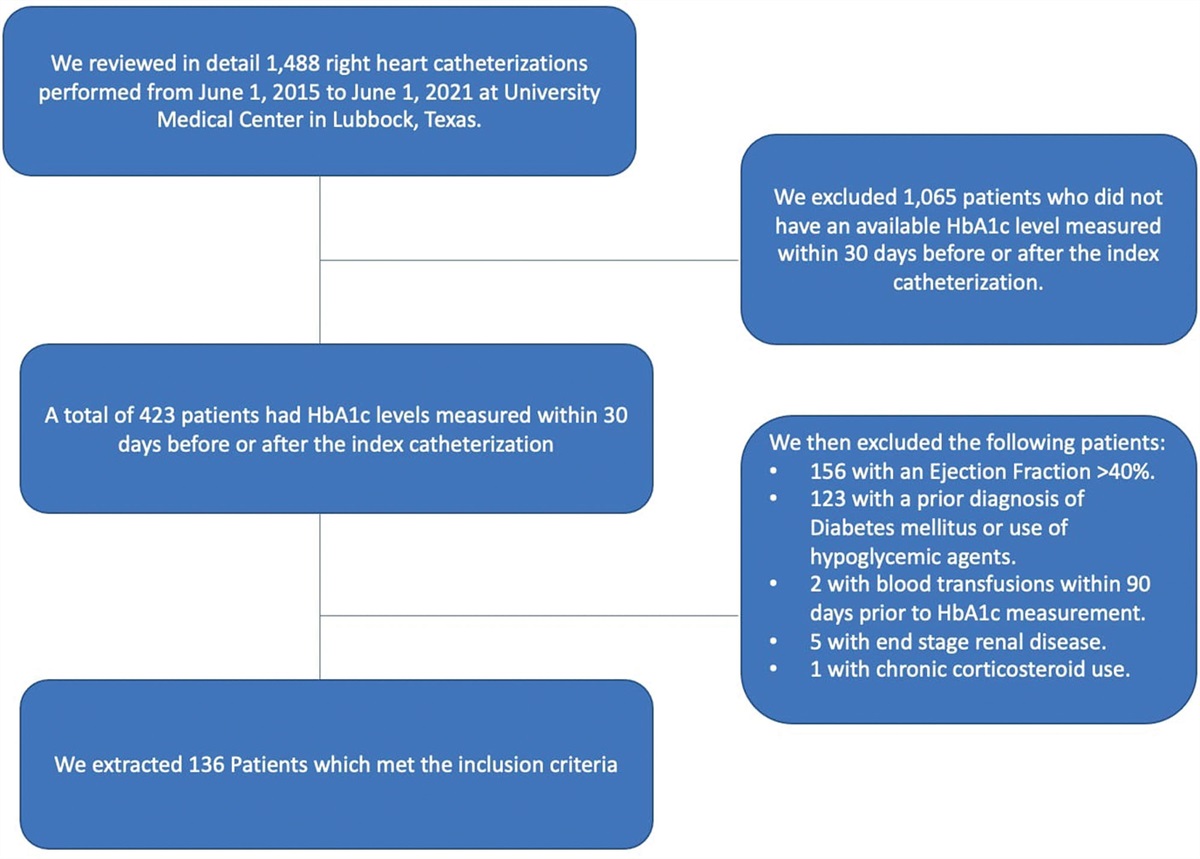
Exclusions were based on questionnaire-derived information regarding prior medical diagnosis. Participants with any history of cardiovascular disease or event were excluded from the study. Participants with documented diagnosis of T2D, and those using renin–angiotensin system inhibitors and beta-blockers, or any medication for managing hypertension or cholesterol were systematically removed from the dataset. Data cleaning process included the removal of incomplete records pertaining to NT-proBNP and essential demographic factors such as age, gender, and BMI. Outliers identified within NT-proBNP and HOMA-IR variables were meticulously identified and removed. The 15th and 85th quantiles of HOMA-IR were selected to ensure a clear categorization of individuals as insulin-sensitive and insulin-resistant, respectively. This selection prevents any ambiguity or placement within the ‘gray zone’, which encompasses individuals near the cutoff range for insulin-sensitive and insulin-resistant classification. Three hundred and ninety-three participants were included in the final analysis.
MetabolomicsUntargeted metabolomics of serum samples from all subjects was performed according to established techniques [15]. Metabolite measurement was performed via Waters ACQUITY ultra-performance liquid chromatography UPLC (Waters Corporation, Milford, Massachusetts, USA) and a Thermo Scientific Q-Exactive high resolution/accurate mass spectrometer (Thermo Fisher Scientific, Inc., Waltham, Massachusetts, USA) interfaced with a heated electrospray ionization (HESI-II) source and Orbitrap mass analyzer operated at 35 000 mass resolution. Compounds were identified by matching with existing library entries of purified standards of over 3300 compounds before being assigned to various categories according to their sources. Quality controls and internal standards are previously described [16]. To track the consistency and replicability of the method over time, quality control samples were employed. To reduce variability and guarantee sample integrity, preanalytical sample handling procedures, such as sample collection, storage, and preparation, followed a standardized methodology.
Statistical analysisThe metabolomics data were inverse rank normalized. Metabolites/pathways associated with antioxidant activity [glutathione metabolism, methionine, cysteine, S‐adenosylmethionine (SAM) and taurine, gamma-glutamyl amino acids, and vitamins (A, C, and E)] were selected for analysis. Principal component analysis was performed to derive a global view of the data. R version 4.2.1 (R Foundation for Statistical Computing, Vienna, Austria) was used to perform linear models for NT-proBNP (as the response variable) versus each metabolite (as the explanatory variables). The model also included the following confounders: age, gender, and BMI. Nominal P-values were penalized using multiple testing correction methods (false discovery rate, FDR). FDR < 0.05 was considered statistically significant. Spearman’s correlation was performed between NT-proBNP and clinical measurements. Statistical significance was determined by P-value <0.01.
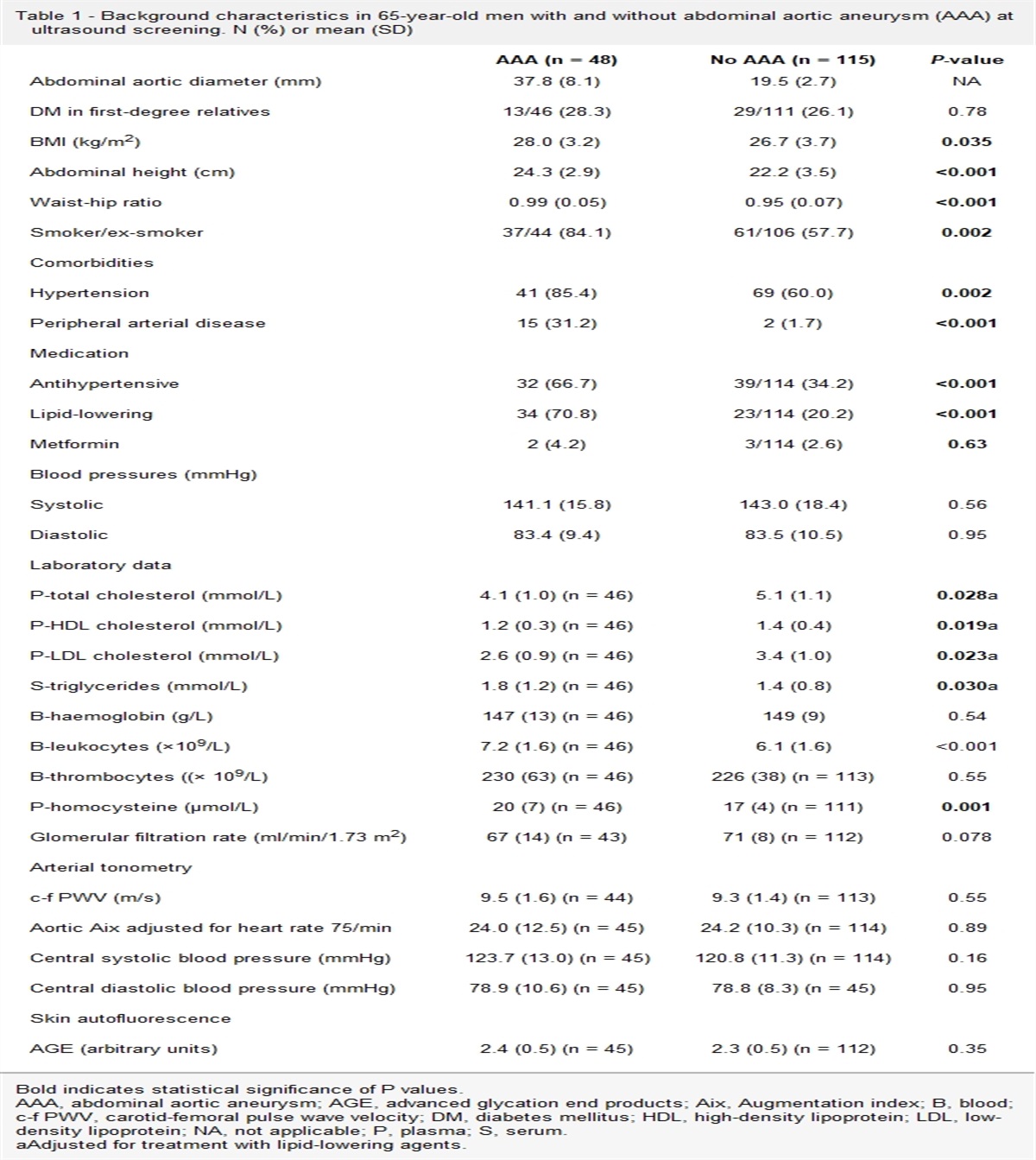
Results General characteristics of participantsThree hundred ninety-three healthy individuals were included in the study. One hundred and eighty-seven were insulin sensitive, and 206 were insulin resistant. Significant differences (Table 1) were shown in BMI, systolic blood pressure, diastolic blood pressure, pulse rate, HOMA-IR, fasting glucose, HbA1C, insulin, C-peptide, total cholesterol, HDL-cholesterol,low density lipoprotein (LDL)-cholesterol, triglyceride, albumin, alanine transaminase, aspartate aminotransferase, gamma-glutamyl transferase (GGT), creatinine, uric acid, hematocrit, and hemoglobin.
Table 1 - General characteristics of participants Test Variable Total (N = 393) Insulin sensitive (N = 187) Insulin resistant (N = 206) Insulin sensitive vs insulin resistantData are presented as mean ± SD or median I(QR) based on the results of the normality test (Shapiro Wilks’). Student’s t-test and Mann–Whitney U test were performed accordingly. Nominal variables were compared using the Chi-square test.
ALT, alanine transaminase; AST, aspartate aminotransferase; GGT, gamma-glutamyl transferase; HDL, high density lipoprotein; HOMA-IR, homeostatic model assessment of insulin resistance; IQR, interquartile range; LDL, low density lipoprotein; TSH, thyroid stimulating hormone.
Spearman correlation was used to assess the relationship between NT-proBNP and clinical traits in the cohort (Fig. 1). Positive correlation was found with HDL. Negative correlation includes, among other clinical parameters, hematocrit, hemoglobin, and GGT.
 Fig. 1: Network of significant correlation between clinical traits and NT-proBNP in the study group. Red/blue reflects the positive/negative Spearman’s ρ, respectively and the thickness of the line depicts the weight of association. (See Supplementary Table 1, Supplemental Digital Content 1, https://links.lww.com/CAEN/A59 for spearman’s ρ and P-values).Effects of antioxidant metabolites on NT-proBNP in the general cohort
Fig. 1: Network of significant correlation between clinical traits and NT-proBNP in the study group. Red/blue reflects the positive/negative Spearman’s ρ, respectively and the thickness of the line depicts the weight of association. (See Supplementary Table 1, Supplemental Digital Content 1, https://links.lww.com/CAEN/A59 for spearman’s ρ and P-values).Effects of antioxidant metabolites on NT-proBNP in the general cohort
A linear model was used to assess the effect of antioxidant-associated metabolites on NT-proBNP level after adjusting for age, gender, and BMI (Table 2). Data suggested a possible decrease in NT-proBNP level with increase of glutamate, while it showed an increase in gamma-glutamylglutamine, S-methylcysteine, cysteine-glutathione disulfide, gamma-glutamylcitrulline, 5-oxoproline, S-methylcysteine sulfoxide methionine, and sulfone gamma-glutamylglycine. Significant correlations are also illustrated in supplementary (Supplementary Figure 1, Supplemental Digital Content 1, https://links.lww.com/CAEN/A59).
Table 2 - Metabolites associated with NT-proBNP after adjusting for age, gender, and BMI. Estimate reflects the effect of the following metabolites on NT-proBNP Metabolite Super pathway Estimate SE P-value FDR Gamma-glutamylglutamine Gamma-glutamyl amino acid 0.170 0.039 1.78 × 10−05 0.001 S-methylcysteine Methionine, cysteine, SAM, and taurine metabolism 0.151 0.036 2.98 × 10−05 0.001 Glutamate Glutamate metabolism −0.161 0.039 4.93 × 10−05 0.001 Cysteine-glutathione disulfide Glutathione metabolism 0.144 0.037 1.25 × 10−04 0.002 Gamma-glutamylcitrulline* Gamma-glutamyl amino acid 0.148 0.040 2.70 × 10−04 0.004 Pyroglutamine* Glutamate metabolism 0.171 0.050 7.39 × 10−04 0.009 S-methylcysteine sulfoxide Methionine, cysteine, SAM, and taurine metabolism 0.110 0.038 3.53 × 10−03 0.035 Gamma-glutamylglycine Gamma-glutamyl amino acid 0.111 0.038 3.90 × 10−03 0.035 Methionine sulfone Methionine, cysteine, SAM, and taurine metabolism 0.107 0.040 7.31 × 10−03 0.058*Indicates a compound that has not been officially confirmed based on a standard but that Metabolon is confident in its identity.
FDR, false discovery rate; SAM, S‐adenosylmethionine.
In order to investigate whether of NT-proBNP levels are affected by insulin resistance status, its levels were compared between insulin sensitive and insulin resistant. Figure 2 shows significantly lower levels of NT-proBNP in insulin-resistant individuals when compared to their insulin-sensitive counterparts. In order to determine metabolites associated with of NT-proBNP in relation to insulin resistance, metabolites differentiating insulin sensitive and insulin resistant were determined. Figure 3 shows higher levels of 5-oxoproline, gamma-glutamylglutamine, gamma-glutamylcitrulline, cysteine-glutathione disulfide, methionine sulfone, S-methyl cysteine (SMC), and SMC sulfoxide in insulin sensitive, whereas methionine, cysteine, cystathionine, glutamate, S-adenosyl-homocysteine, cysteinyl glycine, methionine sulfoxide, and many gamma glutamyl amino acids were higher in insulin resistant.
 Fig. 2:
Fig. 2: Comparing the NT-proBNP level between insulin-sensitive and insulin-resistant individuals in the study group. [****P-value <0.0001].
 Fig. 3: Comparing the metabolite difference between insulin-sensitive and insulin-resistant individuals, FDR https://links.lww.com/CAEN/A59 for estimate values, nominal, and FDR significance).Discussion
Fig. 3: Comparing the metabolite difference between insulin-sensitive and insulin-resistant individuals, FDR https://links.lww.com/CAEN/A59 for estimate values, nominal, and FDR significance).Discussion
Cardiac dysfunction remains a serious global public health problem, and its diagnosis is extremely important. N-terminal fragment brain natriuretic peptides are frequently used in the diagnosis of congestive heart failure [17]. Heart failure is characterized by an exhaustion of the innate antioxidant defense mechanism, leading to an increase in oxidative stress [18]. Metabolomics is now considered an irreplaceable tool in understanding biological functions, by providing insights into the metabolic changes and biomarkers associated with a particular disease or condition. Here, we investigated the possible association between metabolites possessing antioxidant activity, and NT-proBNP in a cohort of healthy individuals; then, we dichotomized our cohort into insulin sensitive and insulin resistant, and compared the levels of these metabolites along with the levels of NT-proBNP between these two groups.
NT-proBNP and clinical traitsOur results showed a significant and inverse association between NT-proBNP with hemoglobin and hematocrit. This is in line with several studies that have found lower hemoglobin levels, often indicative of anemia, are associated with higher levels of NT-proBNP [19–21]. Given that our cohort includes only healthy participants, this suggests that anemia itself can influence NT-proBNP levels, independent of heart failure.
A direct positive association was shown between NT-proBNP and HDL cholesterol. This relation was inversed but less pronounced between NT-proBNP and LDL cholesterol, total cholesterol, and triglyceride. This suggests that an increase in blood NT-proBNP is associated with a beneficial lipide profile. While our findings contradict Zhu et al. [22], they are consistent with Masuch et al. [23], and Birukov et al. [24]. In fact, NT-proBNP has been found to be associated with lipid metabolism, and suggesting a positive role on circulating lipid levels [25]. More studies, however, are needed to extend our understanding of the metabolic properties of NT-proBNP.
NT-proBNP and antioxidant metabolites in the general cohortOxidative stress is a critical hallmark in the pathophysiological cardiac dysfunction in heart failure [1,26]. Glutathione is a tripeptide that plays an important role as antioxidant in the cardiovascular system by counteracting the deleterious effects of ROS and maintaining redox homeostasis [27,28]. Our emerging data showed that increased levels of NT-proBNP was associated with an increase in 5-oxoproline and gamma-glutamylglutamine, and a decrease in glutamate, which may ultimately lead to a reduction in glutathione biosynthesis. Indeed, the enzyme GGT catalyzes the hydrolysis of gamma-glutamyl bonds to form 5-oxoproline, and the enzyme 5-oxoprolinase hydrolyzes 5-oxoproline into glutamate to reconstitute glutathione (Fig. 4). Interestingly, an accumulation in 5-oxoproline owing to downregulation of 5-oxoprolinase or an increased in GGT activity, was shown to be an important factor in heart failure [29,30]. Relatedly, Sourdon et al. showed that disturbing 5-oxoprolinase activity can trigger what they called a redox storm in the ischemic heart [31]. Moreover, many studies suggest participation of GGT in the pathophysiology of cardiovascular diseases [32,33].
 Fig. 4:
Fig. 4: Metabolic antioxidant pathways significantly associated with NT-proBNP in this study.
Cumulative evidence attests that altered GSH cycling can trigger cardiac oxidative stress. In heart failure, a significant decrease in glutathione levels has been observed [30,34]. Kobayashi et al. showed that an increased vulnerability of mitochondria to oxidative damage caused by a reduction in GSH, was associated with a higher susceptibility of myocardial injury in mice [35].
Our findings were further corroborated by the direct significant association between NT-ProBNP levels with SMC and its metabolite SMC sulfoxide. The increase in the levels of these metabolites could indicate a divergence in cysteine metabolism away from glutathione synthesis due to the low availability of glutamate (Fig. 4). Relatedly, El-Magd et al. [36] demonstrated that high doses of SMC have been found to result in a significant increase in serum cardiac injury biomarkers. Further experiments, however, are required to check the exact involvement of SMC in cardiac function.
Our emerging data also showed a direct and significant association between NT-ProBNP levels with cysteine-glutathione disulfide levels. The accumulation of cysteine-glutathione disulfide can indicate an increased level of oxidative stress in the body [37], which leads to formation of glutathione mixed disulfides with protein thiol groups, leading to a process called S-glutathionylation [38]. Intriguingly, S-glutathionylation is emerging as a critical signaling mechanism in cardiovascular diseases [39,40]. Additionally, S-glutathionylation of cysteine is catalyzed by glutathione S-transferase P1 [41], an enzyme strongly associated with heart failure [42]. More research, however, is needed to understand the role of S-glutathionylation in cardiac disorders, as well as its potential use in therapies and as a biomarker.
The increase of NT-ProBNP associated with the increase in methionine sulfone can also be explained by an increase in ROS overwhelming the cellular antioxidant defense system. Methionine is oxidized to methionine sulfoxide; this oxidation can be reversed by enzymes in the methionine sulfoxide reductase family which scavenge ROS; however, the further oxidation of methionine sulfoxide to form methionine sulfone is irreversible [43]. Strikingly, methionine sulfoxide reductase A was shown to protect the heart from ischemia-reperfusion injury [44], the latter being a significant factor in the development of heart failure [45].
NT-proBNP and antioxidant metabolites in insulin sensitivity and insulin resistanceOur results showed significantly lower levels of NT-ProBNP in insulin resistant participants when compared to insulin-sensitive counterparts. This is in line with many studies [12,46–48], which showed that insulin resistance, is generally associated with reduced NT-proBNP levels.
Interestingly, our results showed that all metabolites positively associated with NT-ProBNP in the general cohort, were lower in the insulin resistant group when compared to insulin-sensitive counterpart. This includes 5-oxoproline, gamma-glutamylglutamine, gamma-glutamylcitrulline, cysteine-glutathione disulfide, methionine sulfone, SMC, and SMC sulfoxide. Moreover, glutamate, negatively associated with NT-ProBNP in the general cohort, was higher in insulin resistant group when compared to insulin sensitive. This shows that the decrease of NT-ProBNP observed in insulin resistant group may be associated, at least in part, with the change of these metabolites.
Besides glutamate, the decrease of NT-ProBNP in the insulin resistant group, was also accompanied by an increase of cystathionine, cysteine, S-adenosyl-homocysteine, cysteinyl glycine, methionine, and many gamma-glutamyl amino acids. This may indicate a potential indirect enhancement in the glutathione cycle via the transsulfuration pathway. Indeed, cysteine is the immediate precursor of glutathione, and the transsulfuration pathway is the only route for biosynthesis of cysteine from homocysteine via cystathionine [49] (Fig. 4). Studies have shown that an activation of the transsulfuration pathway significantly contributes to an increase in glutathione biosynthesis [50]. The increase in glutathione synthesis in insulin resistant could be a compensatory mechanism to reduce oxidative stress and improve insulin signaling [51]. Moreover, recent research has shown that glutathione supplementation can improve insulin sensitivity [52]. Conversely, the insulin-sensitive group featuring higher NT-ProBNP exhibited higher levels of 2-amino butyrate and 2-hydroxy butyrate. A study on murine cardiomyopathy model showed that increased ROS are accompanied by 2-amino butyrate accumulation and compensatory maintenance of myocardial GSH levels [53]. Remarkably, α-hydroxybutyrate dehydrogenase (α-HBDH) is a marker of cell death reflecting myocardial injury [54]; the latter condition was shown to be associated with an increase in NT-proBNP [55].
Taken together, our results provide evidence that an increase in NT-proBNP levels could be ascribed, at least partially, to an imbalance in glutathione cycle and accumulation of ROS; and the reduction of NT-proBNP observed in insulin-resistant individuals, could be a result of a direct or indirect restoration of glutathione biosynthesis.
ConclusionOur study provides the research community with a wealth of antioxidant metabolites associated with NT-proBNP in insulin resistance. These metabolites are directly or indirectly involved in the glutathione metabolism. We believe that the compensation mechanisms demonstrated in insulin resistance, which target the reduction of oxidative stress and enhancement of insulin signaling, may play a role in the observed decrease in NT-proBNP levels. Therefore, these metabolites can be of clinical relevance to provide valuable information about the heart’s functional status. Additionally, the study could advance our understanding of the interplay between oxidative stress during insulin resistance and cardiovascular risk, which could lead to novel therapeutic approaches for managing cardiovascular diseases. We acknowledge limitations in our study; while glutathione data is very useful to our interpretation, its measurement in nontargeted metabolomics is not typically performed due to the nature of the nontargeted approach and the specific challenges associated with accurately measuring glutathione levels. An additional limitation is the absence of echocardiographic data for participants. Another limitation is the measurement of the levels of NT-proBNP in healthy individuals; since reference ranges for NT-proBNP in healthy individuals are not well-established and can vary depending on the population studied and it can be affected by various extracardiac factors. Further studies are warranted to investigate the relevant usage of these metabolites in clinical practice, and to understand how these differences in oxidative stress markers translate into variations in NT-proBNP values.
AcknowledgementsThe authors would like to acknowledge Qatar Biobank for providing the data.
This research was funded by Qatar National Research Fund (QNRF), grant number NPRP13S-1230-190008.
K.N. wrote the article, N.A. wrote the article and supported the statistical analysis, and M.A.E. supervised the work. All authors have read and agreed to the published version of the manuscript.
The study was approved by the Institutional Review Boards of the Qatar Biobank (QF-QBB-RES-ACC-00125).
Informed consent was obtained from all participants involved in the study.
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Conflicts of interestThere are no conflicts of interest.
References 1. Tsutsui H, Kinugawa S, Matsushima S. Oxidative stress and heart failure. Am J Physiol Heart Circ Physiol 2011; 301:H2181–H2190. 2. Tangvarasittichai S. Oxidative stress, insulin resistance, dyslipidemia and type 2 diabetes mellitus. World J Diabetes 2015; 6:456–480. 3. Ormazabal V, Nair S, Elfeky O, Aguayo C, Salomon C, Zuniga FA. Association between insulin resistance and the development of cardiovascular disease. Cardiovasc Diabetol 2018; 17:122. 4. Federico C. Natriuretic peptide system and cardiovascular disease. Heart Views 2010; 11:10–15. 5. Sarzani R, Allevi M, Di Pentima C, Schiavi P, Spannella F, Giulietti F. Role of cardiac natriuretic peptides in heart structure and function. Int J Mol Sci 2022; 23:14415. 6. Cao Z, Jia Y, Zhu B. BNP and NT-proBNP as diagnostic biomarkers for cardiac dysfunction in both clinical and forensic medicine. Int J Mol Sci 2019; 20:1820. doi: 10.3390/ijms20081820. 7. Szczurek W, Szyguła-Jurkiewicz B. Oxidative stress and inflammatory markers - the future of heart failure diagnostics? Kardiochir Torakochirurgia Pol 2015; 12:145–149. 8. Suzuki K, Ishii J, Kitagawa F, Kuno A, Kusuhara Y, Ochiai J, et al. Association of serum carotenoid levels with N-terminal pro-brain-type natriuretic peptide: a cross-sectional study in Japan. J Epidemiol 2013; 23:163–168. 9. Meng XM, Kang SX, Li J, Zhang HT, Li M. Clinical significance of N-terminal natriuretic peptide combined with inflammatory factors, oxidative stress factors and blood lipid detection in elderly patients with type-2 diabetes complicated with coronary heart disease. Pak J Med Sci 2022; 38:1310–1315. 10. Chen Q, Qi S, Hocum-Stone L, Lesnefsky E, Kelly RF, McFalls EO. Preventing myocardial injury following non-cardiac surgery: a potential role for preoperative antioxidant therapy with ubiquinone. Antioxidants (Basel) 2021; 10:276. 11. Fu S, Ping P, Zhu Q, Ye P, Luo L. Brain natriuretic peptide and its biochemical, analytical, and clinical issues in heart failure: a narrative review. Front Physiol 2018; 9:692. 12. Echouffo-Tcheugui JB, Zhang S, McEvoy JW, Juraschek SP, Fang M, Ndumele CE, et al. Insulin resistance and N-terminal pro-B-type natriuretic peptide among healthy adults. JAMA Cardiology 2023; 8:989–995. 13. Hollstein T, Schlicht K, Krause L, Hagen S, Rohmann N, Schulte DM, et al. Effect of various weight loss interventions on serum NT-proBNP concentration in severe obese subjects without clinical manifest heart failure. Sci Rep 2021; 11:10096. 14. Albert CL, Tang WHW. Metabolic biomarkers in heart failure. Heart Fail Clin 2018; 14:109–118. 15. Al-Khelaifi F, Diboun I, Donati F, Botrè F, Alsayrafi M, Georgakopoulos C, et al. A pilot study comparing the metabolic profiles of elite-level athletes from different sporting disciplines. Sports Med Open 2018; 4:2. 16. Evans A, Bridgewater B, Liu Q, Mitchell M, Robinson R, Dai H, et al. High resolution mass spectrometry improves data quantity and quality as compared to unit mass resolution mass spectrometry in high-throughput profiling metabolomics. Metabolomics 2014; 4:2. doi: 10.4172/2153-0769.1000132. 17. Maries L, Manitiu I. Diagnostic and prognostic values of B-type natriuretic peptides (BNP) and N-terminal fragment brain natriuretic peptides (NT-pro-BNP). Cardiovasc J Afr 2013; 24:286–289. 18. van der Pol A, van Gilst WH, Voors AA, van der Meer P. Treating oxidative stress in heart failure: past, present and future. Eur J Heart Fail 2019; 21:425–435. 19. Tirmenstajn-Jankovic B, Dimkovic N, Perunicic-Pekovic G, Radojicic Z, Bastac D, Zikic S, et al. Anemia is independently associated with NT-proBNP levels in asymptomatic predialysis patients with chronic kidney disease. Hippokratia 2013; 17:307–312. 20. Willis MS, Lee ES, Grenache DG. Effect of anemia on plasma concentrations of NT-proBNP. Clin Chim Acta 2005; 358:175–181. 21. Lelli D, Antonelli Incalzi R, Pedone C. Hemoglobin concentration influences N-terminal pro B-type natriuretic peptide levels in hospitalized older adults with and without heart failure. J Am Geriatr Soc 2017; 65:2369–2373.
Comments (0)