
Remember me







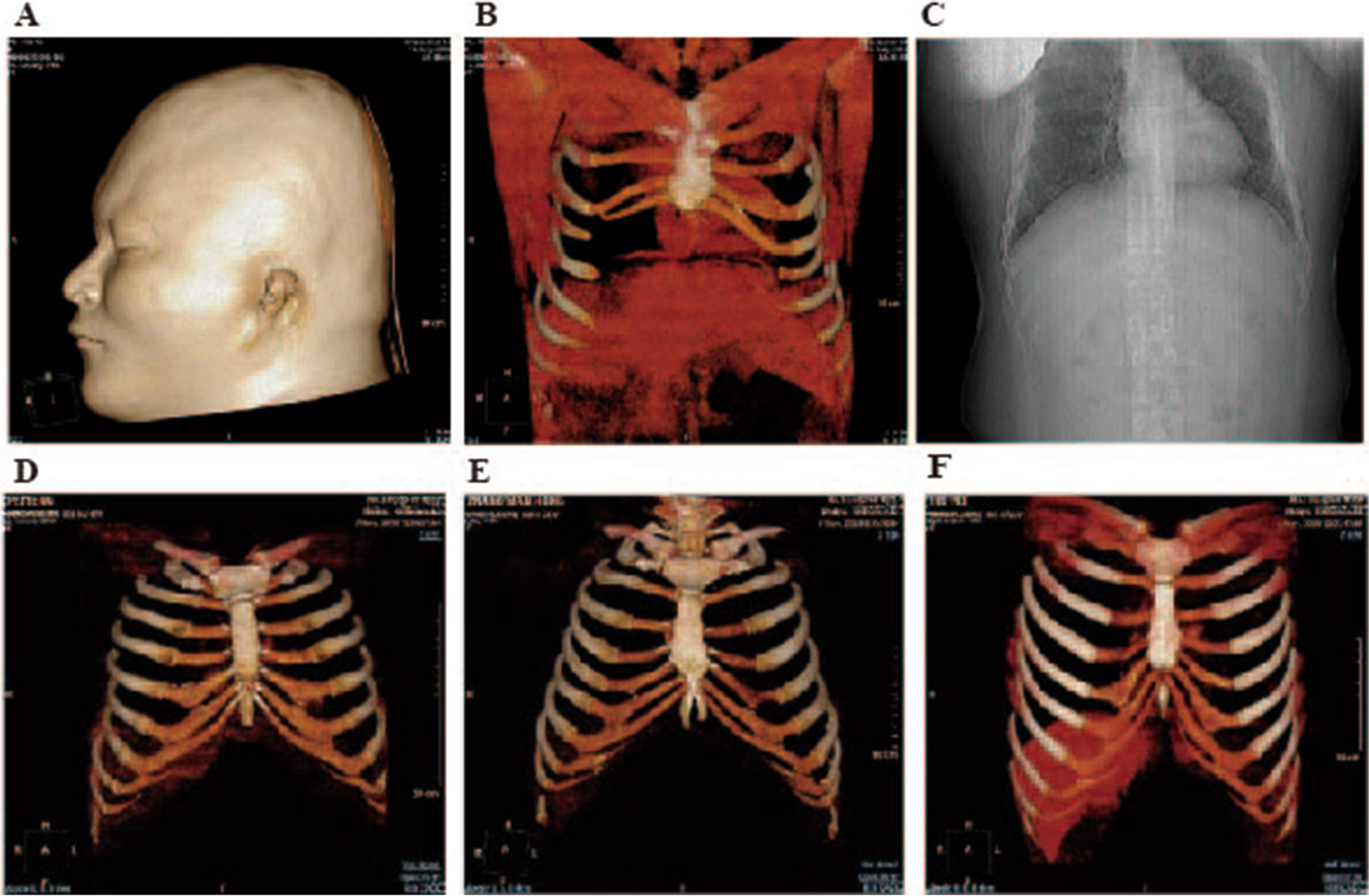
Microtia is one of the leading congenital malformations in the plastic surgery field, it brings harm to children's psychology, and puts pressure on their families and increases society's burden, experts are attaching increasing importance to the exploration of microtia. One major obstacle in treating microtia is that the underlying mechanism is complicated and not clearly understood. Owing to the fact that auricular cartilage originates from NCCs [10], therefore, previous reports about microtia mainly focused on the development of NCCs. In this study, we further investigated the etiology of microtia accompanied by costal chondrodysplasia and sternal bone deformity by bioinformatic tools. WES and label-free analysis were conducted between the family members. Interestingly enough, both WES and proteomic results indicated that metabolic program, associated immune responses, and inflammatory and cellular redox homeostasis may play a pathological or at least a modulating role in microtia, and this has never been reported before.
The candidate genes SOD3 and PLG by ACMG classification overlap with the proteomic results. PLG not only regulates coagulation and complement pathway, it is also responsible for neutrophil apoptosis and efferocytosis in inflammation, and functions as apolipoprotein binding [11], whereas in our case, it is downregulated in microtia patients when compared to his mother but upregulated than his father. It is widely believed that excessive reactive oxygen species (ROS) can lead to oxidative stress and DNA damage which can decrease MSCs self-renewing and multidirectional differentiation [12]. SOD3 protects cells from the toxic effect of RO intermediates by converting superoxide radicals into hydrogen peroxide and oxygen. Nightingale et al. found that decreased SOD3 level following BMSCs chondrogenesis [13]. Gavriilidis et al. knocked down SOD2 in chondrocytes, resulting in increased ROS levels, mitochondrial DNA strand breaks and decreased antioxidant capacity, and this may be a potential contributor to osteoarthritis [14].
The variant identified in the PRKDC gene is predicted to be pathogenic according to ACMG, it can recognize and repair DNA double-strand breaks [15], and interacts with autoimmune regulator (AIRE) to regulate B cells and natural killer (NK) cells, causing inflammation and immune dysregulation, and downstream S100A8 expression [16]. Plus, PRKDC is also involved in nervous system development [17]. Although S100A8 does not show significance in WES, it is upregulated in proteomics results. S100A8 is a calcium- and zinc-binding protein that plays a prominent role in the regulation of inflammatory processes and immune response. S100A8/A9 promotes cell death via autophagy and apoptosis and this occurs through the crosstalk of mitochondria and lysosomes via ROS [18]. The S100 family is closely related to chondrogenesis [19], and S100A8 and S100A9 up-regulate inflammatory cytokines through TLR-4 And MMP-1, -3, -9, and -13 to destroy bone [20].
TASP1 encodes taspase 1(threonine aspartase 1, TASP1). TASP1 proenzyme intramolecularly proteolyzes and generates an active N-terminal 28 kDa α subunit and a 22 kDa C-terminal β subunit heterodimer whereas the β subunit is the active site for cleavage activity [21]. TASP1 cleaves various nuclear factors after an aspartate with substrates including the histone-methyltransferase mixed lineage leukemia 1 (MLL1/KMT2B), transcription factor IIA (TFIIAα-β/GTF2A1), TFIIA-like factor (ALFα-β) and so on [22]. TASP1 activateing MML1 is crucial for HOX and cyclin genes expression that participate in body segmentation and cell proliferation [23] and TASP1 cleaving TFIIAα-β make it more susceptible to degradation impedes its transcriptional activity in embryonal cell proliferation and morphogenesis. Tasp1-/- mice show craniofacial malformations [24] and facial features containing low-set ears [25] and TASP1 mutation has been reported in complex syndromes manifesting with facial and skeletal abnormalities [26]. As mitochondrial redox status can affect SAM level to modify histone methylation [27], and studies have identified SET/MLL as redox sensitive units of the H3K4me3 [28]. The relationship between Taspases and oxidative stress-mediated alterations in DNA/histone methylation and other post-translational modifications is worth to be explored in future studies.
The enrichment analysis of differently expressed genes (DEGs) revealed that mutated genes are majorly enriched in the cellular metabolic process, embryonic development process, oxidative stress and cytoplasmic vesicles, which are concordant with the results in proteomics analysis. It may reflect that metabolism may play an important regulating role in the pathogenesis of microtia. Expression differences could be a sign of the biochemical or physiological changes that might lead to microtia. Many researches have proved that immunometabolism is closely related to the development of diseases [29]. Oxidized low-density lipoprotein can trigger strong proinflammatory responses that can potentially contribute to the development of atherosclerosis [30].
What gives us implications in our study is the upregulated proteins in three groups are closely related to the immune system and oxidative stress. The highest change of protein is ORM2, which is reported to modulate the activity of the immune response and the complement cascades pathway [31]. ORM2 is at the same time a suppressor of Fe-related ROS [32]. SAA1 is also a major acute-phase inflammatory protein [33], it contributes to bone and cartilage destruction [34]. IGHD as its name implies, mediate the effector phase of humoral immunity [35]. Polymeric immunoglobulin receptor (PIGR) specifically binds to J-chain-containing secretory IgM and IgA, therefore, it is a critical protein in the mucosal immunity [36]. Vertebrate haemoglobin, a heterotetramer of paralogous α- and β-subunits that mediate respiratory oxygen transport and exchange [37], and can be cleared by binding to the haptoglobin (HP), in this way to scavenging HBB-mediated toxic effects caused by free radical and participates in the complementary pathways [38].
When comparing the proteomics results to the WES SNPs genes, there are overlapped results: MASP2 and FCN2, which are all belong to key proteins of the lectin pathway of complement. Ficolin 2 is the initiator and their N-terminal collagen-like domain interacts and forms complexes with the MASPs, which leads to the C4 and C2 cleavage and thus activation of the downstream complement cascade [39]. In our results, FCN2 is at least fivefold higher, and the variation is heterogenous in his mother. MASP2 is also at least fivefold higher and the variation is heterogenous in the patient’s sister and father, both are in the same trend with the proteomics results. It is reported that a higher level of ficolin is related to autoimmune disorders [40]. There have been reports about complement-associated and prostaglandin-dependent bone metabolism and physiologic remodeling [41], such as axial spondyloarthritis [42]. It is also believed that complement activation can release cytokines, proteases, and ROS, thereby contributing to inflammation and generates deleterious effects [41]. But so far, none of these molecules have been reported in the pathology of microtia.
Cadherin-13 (CDH13) that repeat in the groups A vs. B and A vs. D is an important adhesion molecule that mediates cell–cell adhesion in neural tube development [43], while there is a report about CDH13 and CRTAC expression in the limb mesenchyme control contrarily proprioceptor identity [44]. Coincidently, CRTAC is downregulated in our WES result. Cdh13 protects against cardiac stress through its association with adiponectin through the AMPK signaling pathway in mice [45]. What is more, research has proved that, Cdh13 serves as an anchor for tethering of adiponectin to M2 macrophages and can promote the cell proliferation by activation of Akt [46]. In our proteomic results, CDH13, adiponectin and CRTAC are all downregulated, which may imply the disruption of the cell proliferation and neural tube development, and inflammation may participate in the malfunction.
Besides, the commonly dysregulated genes were significantly enriched in GO terms in Fig. 3 and Table 9 shows that most proteins are significantly enriched in lipid metabolic process, peroxidase activity, and the hemoglobin related functions. LEPR+ MSCs give rise to most bone and adipocytes in adult bone marrow and can form bony ossicles that support hematopoiesis in vivo [47]. ApoE is involved in lipoprotein metabolism and ApoE -/- mice showed intervertebral disc degeneration partially due to increased matrix metalloproteinase (MMP) -3, -9, and -13 levels [48]. In addition, Farnaghi et al. found that hypercholesterolemia caused the degeneration of articular disc cartilage in osteoarthritis by the production of mitochondrion-derived oxidative stress [49]. Evidence shows that there is a link between oxidative stress and cartilage degradation.
What is more, emerging evidence has shown that cell metabolism influences gene expression by modifying the epigenome, which can regulate stem cell pluripotency, differentiation and somatic cell reprogramming [50]. ALDOB participates in fructose and other central carbon metabolism by reversibly cleaving fructose 1,6-diphosphate into glyceraldehyde 3-phosphate (G3P) and dihydroxyacetone phosphate (DHAP) [51]. Lipogenesis requires oxidative phosphorylation to provide a large amount of energy for lipid storage, while chondrocytes mainly rely on glycolysis for energy supply [52]. We hypothesize that reduced ALDOB protein levels in microtia patients may lead to decreased glycolysis and decreased MSC’s chondrogenic ability in some way which would be studied in future studies.
In label-free proteomics analysis, ALDOB protein is 5.41239 times higher in the patient compared to his sister(B) and 3.87038 times higher when compared to the mother in the A/C group, CDH13 and ADIPOQ are 0.35024 and 0.26241 times in A vs. B group respectively and show the same trend in A vs. D group by 0.31112 and 0.29045 time respectively. This is coherent with the result in IHC that CDH13 and ADIPOQ are lower in microtia patients. But the real-time PCR of other microtia patients shows that mRNA levels of ALDOB, ADIPOQ, CDH13, and TASP1 are all higher in the microtia group while IHC results show all of the four proteins are significantly lower when compared to the control group as shown in Figs. 7 and 8. This is probably due to the translational modifications of the mRNAs and shall be further explored as a candidate disease mechanism.
Fig. 8
Graphical Abstract. A graphical overview of this study
The collective findings provide novel insights into the pathogenesis of congenital microtia-atresia. It can be seen that the screened proteins are mainly involved in lipid metabolism, immunology, and inflammation process, and our in vitro verifications have proved that ROS do exist in microtia patients, as some fault programmed gene expression and misregulated microenvironment during the embryonic stage could be kept in the entirety and outer ear keeps on developing after birth [53], therefore these faults may affect ear length, width, prominence and degree of auricular development. The relationships between oxidative stress and these proteins in cancers or inflammatory processes have been widely studied [29], but the relationship between oxidative stress and chondrogenic defect of microtia has not been thoroughly realized. Alterations of the redox state in microtia may provide new insight into pathogenic mechanisms and refine therapeutic strategy, with the ultimate goal of improving the quality of life for the patients.
Potential limitationsThere are also some limits of proteomic analyses as protein may not fully reflect the genetic variants and can only show the pathology of micro-environmental effect in this case. To overcome this limitation, we use the whole-exome sequencing to find out the possible variations in microtia at the genome level. However, the exome region constitutes only 1–2% of the entire genome, and WES inevitably leaves out the non-coding mRNA elements and/or intronic or intergenic regulatory regions that may be responsible for the disease phenotypes [2]. What is more, due to individual variances, more microtia patients are needed to further verify our findings. Last but not least, more explorations of how oxidative stress and malfunction of genes related to embryonic development in vivo shall be done in our future studies.
Comments (0)