
Remember me






Insomnia is a highly prevalent disorder that involves persistent difficulties falling or staying asleep and is often associated with excessive daytime sleepiness, fatigue, or other functional impairments [1]. A leading theory suggests that primary insomnia is due in large part to hyperarousal at bedtime, often driven by negative ruminative thoughts, worry, introspection, and other disturbing or arousing pre-sleep cognitions [1]. Notably, this type of self-focused cognition is associated with increased activation of the brain’s default mode network (DMN) [2], a large-scale network that includes the medial prefrontal cortex (MPFC), posterior cingulate/precuneus region, and bilateral regions of the inferior parietal cortex, as well as several less prominent areas [3]. The well-documented association between DMN activation and self-directed cognition has led to a growing body of neuroimaging research examining the activation and resting state functional connectivity (rsFC) patterns of the DMN among patients with insomnia disorders [4,5]. There is now extensive evidence implicating altered functioning of the DMN as a primary feature of insomnia [6].
Virtually all research on DMN connectivity in people with insomnia has focused on associations with subjective scales or clinical symptoms rather than objective measures of sleep using polysomnography (PSG). One exception found that increased within-network connectivity of the DMN was associated with lower sleep efficiency, reduced rapid eye movement (REM) sleep, and longer latency to fall asleep on PSG [7]. However, that study only addressed within-network connectivity and it is quite likely that the DMN interacts with other brain regions, such as the prefrontal cortex and other sensory, motor, and affective processing regions to influence sleep. To our knowledge, no neuroimaging studies have examined the associations of DMN-to-whole brain (i.e. seed-to-voxel) connectivity and subsequent PSG-measured sleep outcomes. Accordingly, we collected rsFC data in the evening just prior to a PSG-monitored overnight laboratory sleep opportunity in a sample of individuals scoring high on measures of insomnia symptoms. The term ‘resting state’ simply refers to the fact that no external stimulation or manipulation occurs during the scan. Thus, the outcome metric is the intrinsic connectivity among brain regions as the participant allows their thoughts to wander without explicit constraints. In this correlational study, we hypothesized that rsFC between the DMN and cortical regions involved in attention, decision-making, and interoceptive awareness prior to bedtime would predict subsequent PSG sleep metrics.
Methods ParticipantsTwenty adults (8 men and 12 women) between the ages of 19 and 39 years (M = 26.9, SD = 6.6) with normal sleep/wake schedules, but scoring above clinical criteria for current insomnia symptoms, completed all study procedures. To be eligible, participants had to score above threshold on at least two of the following three sleep questionnaires (≥15 on the Insomnia Severity Index [8]; ≥6 on the Pittsburgh Sleep Quality Index [9]; ≥11 on the Epworth Sleepiness Scale [10]). Participants were excluded for MRI contraindications, breathing-related disorders, and medical, psychiatric, or neurologic conditions. Other exclusions include travel across time zones within one week of the study, current use of sleep-altering medications or supplements, relevant substance use (excessive caffeine, nicotine, alcohol, illicit drugs), or less than ninth-grade English reading equivalent. The protocol for this project was approved by the Institutional Review Board at the University of Arizona and the U.S. Army Human Research Protections Office. All participants provided written informed consent before enrollment.
Materials and procedureParticipants completed a baseline intake that included the Wechsler Abbreviated Scale of Intelligence-II (IQ) [11], and returned about a week later for a brain scan and overnight sleep assessment. Participants were required to maintain a regular sleep schedule for the week prior to the in-lab stay and to abstain from caffeine-containing products for 48 h before reporting to the study. These procedures were part of a larger study examining the effects of repetitive transcranial magnetic stimulation (rTMS) on sleep and brain function, however, all data reported here were collected during the night following the sham condition (i.e. no participants received any form of active TMS prior to MRI or sleep). At 8:30 p.m. on the evening of the overnight run, after completing a 40-second sham rTMS session, each participant underwent a series of MRI scans including a 6-minute rsFC scan. Upon completion of the scan, each participant was escorted to a private, sound-attenuated, temperature-controlled bedroom where they were allowed to eat a pre-selected meal and were fitted with polysomnographic (PSG) electrodes with a standard 10–20 placement. Lights were turned off at 11:00 p.m. and participants were monitored over an 8-hour sleep period using a Nihon Kohden JE-921 PSG recording system. Data from PSG were subsequently scored using Polysmith software by a trained and certified sleep technician who was blind to participant status. The overnight PSG metrics including number of minutes and percentage of time spent in N1, N2, N3, and rapid eye movement (REM) sleep, as well as total sleep time (TST; i.e., the total number of minutes of time scored as sleep) and sleep efficiency (SE; i.e., the percent of time in bed spent in sleep) were manually scored by the technician using the standard Rechtschaffen and Kales scoring approach [12]. Due to an unanticipated morning disruption during the last hour of sleep for one participant, sleep stage scoring was truncated to the first 7 hours of sleep (i.e. from 11:00 p.m. to 6:00 a.m.) for all participants to insure uniformity in sleep staging.
Neuroimaging methodsNeuroimaging data were collected with a Skyra 3T MRI scanner (Siemens, Erlangen, Germany) with a 32-channel head coil. First, a T1-weighted structural MRI was acquired with a 3D MPRAGE sequence (TR/TE/flip angle = 2.1 s/ 2.33 ms/12°) across 176 sagittal slices (256 × 256) and a slice thickness of 1 mm (voxel size = 1 × 1 × 1 mm3). Resting-state functional MRI (T2*) scans were acquired over 60 transverse slices (2.0 mm thickness; matrix: 128 × 128) using an interleaved sequence (TR/TE/flip angle = 2 s/36 ms/90°; 2.0 × 2.0 voxel size (i.e. with a 10% slice gap, 180 volumes; FOV = 240 mm) over a 6-min acquisition time. To minimize noise in the scanner, all participants wore foam earplugs. Participants were given the following instruction: ‘Please keep your eyes open, focused on the fixation point. Take this time to rest and let your thoughts come and go. Remember to stay as still as possible’.
Resting state connectivity preprocessingThe rsFC data were analyzed using the CONN functional connectivity toolbox (version 21.a; www.nitrc.org/projects/conn). Functional volumes were realigned, unwarped, slice-time corrected, and coregistered with individual T1 images in accordance with standard algorithms. Noise was removed with the aCompCor method [13,14], band-pass filtering (0.01–0.1 Hz), tissue segmentation, and removal of confounding effects through linear regression. Nuisance covariates (i.e. scan-to-scan motion exceeding 0.5 mm; >3 SD mean global signal intensity) were regressed out of the first-level analysis. Functional images were normalized to the Montreal Neurologic Institute stereotaxic space and spatially smoothed (8 mm full width at half maximum), and resliced to 2 × 2 × 2 mm.
Functional Connectivity AnalysisWithin CONN, the four primary nodes of the DMN (i.e. MPFC; left lateral parietal cortex; right lateral parietal cortex; posterior cingulate cortex), were selected as seed regions based on the standard DMN atlas included in the software (Fig. 1a). All four seed regions were weighted equally in the between-sources contrast and analyzed for whole-brain connectivity. These connectivity maps were then correlated separately with each PSG sleep variable, after controlling for age, sex, and IQ, implementing a voxel threshold of P < 0.005 (uncorrected) and a cluster size threshold of P < 0.05 (FDR corrected). This cluster correction approach reduces the probability that the observed correlations between connected regions and subsequent sleep stages are spurious (i.e. reduces Type I error). The outcomes of interest focus on the correlation between overall DMN-region connectivity and individual overnight PSG measures, but the figures also provide plots of the effect sizes for each of the individual associations for each DMN region separately.
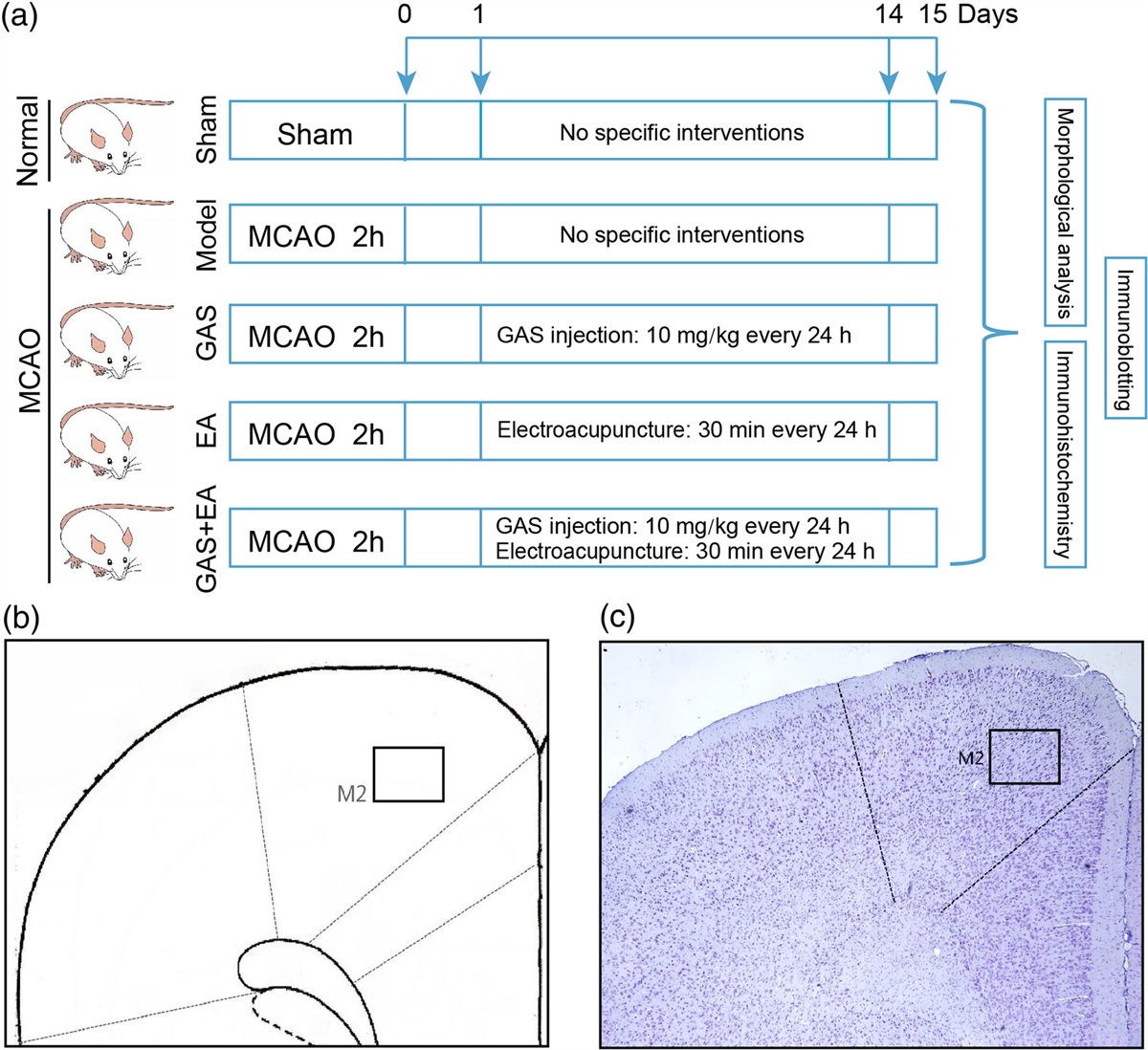
 Fig. 1:
Fig. 1: Regions showing connectivity with the Default Mode Network (DMN) that correlates with each polysomnographically measured sleep stage. The bar graphs show the strength of association for each DMN node separately. (a) Map showing the four primary nodes of the DMN that were used as a combined seed region in the present analysis (green = medial prefrontal cortex node—MPFC; turquoise = left lateral parietal node—LP (L); magenta = right lateral parietal node—LP (R); purple = posterior cingulate/precuneus node (PCC). (b) Connectivity between the DMN and right angular gyrus is associated with fewer minutes of wake during the night. (c) Connectivity between the DMN and the precuneus (PC) was associated with less stage N1 sleep during the night. (d) Connectivity between the DMN and the left orbitofrontal cortex (OFC) was positively correlated with greater rapid eye movement (REM) sleep. (e) Connectivity between the DMN and several regions including the left OFC, left insula, superior frontal gyrus, and right anterior cingulate cortex was associated with greater minutes of total sleep time (TST). (f) Connectivity between the DMN and regions including the left OFC (red = positive correlation) and the pons (blue = negative correlation) were each associated with opposite correlations with sleep efficiency (SE).
Results WakeAs shown in Table 1 and Fig. 1b, greater connectivity between the DMN and the right angular gyrus (AG) was associated with fewer minutes of wake (R2 = 0.79) and lower percentage of wake during the sleep period (R2 = 0.77).
Table 1 - Correlation statistics between each PSG sleep parameter and DMN-to-region functional connectivity Sleep parameter Units Region MNI coordinate Region label T (df = 15) Cluster size Cluster p-FDR Sleep parameter-connectivity correlation Beta weights for DMN nodes MPFC L LP R LP PCC Wake (min) 50, −44, 16 R Ang Gyrus −6.69 721 < 0.001 Negative −0.004 −0.005 −0.006 −0.004 (%) 50, −44, 16 R Ang Gyrus −6.11 616 < 0.001 Negative −0.017 −0.021 −0.027 −0.019 N1 (min) 6, −46, 46 R Precuneus −5.11 499 < 0.001 Negative −0.006 −0.014 −0.017 −0.009 (%) −2, −48, 68 L Precuneus −4.28 187 0.026 Negative −0.013 −0.048 −0.056 −0.033 N2 (min) -- -- -- -- -- -- -- -- -- -- (%) −38, 22, 4 L Insula −5.25 314 < 0.001 Negative −0.018 −0.012 −0.014 −0.013 8, 50, −20 R Vent Med FC 5.42 151 0.046 Positive 0.021 0.008 0.022 0.007 N3 (min) -- -- -- -- -- -- -- -- -- -- (%) -- -- -- -- -- -- -- -- -- -- REM (min) −32, 34, −2 L OFC 7.01 177 0.021 Positive 0.006 0.005 0.003 0.003 (%) −32, 36, −4 L OFC 6.89 164 0.032 Positive 0.024 0.025 0.016 0.015 TST (min) −36, 16, −8 L Insula 5.24 273 0.002 Positive 0.200 0.142 0.166 0.129 −16, 24, −26 L OFC 5.97 179 0.015 Positive 0.197 0.154 0.091 0.105 8, 6, 68 L/R SFG 6.65 161 0.017 Positive 0.144 0.133 0.171 0.136 8, 48, 26 R SFG/ACC 5.67 123 0.046 Positive 0.162 0.115 0.147 0.167 SE (%) 16, −14, −40 Brainstem/Pons −7.42 267 0.003 Negative −0.804 −0.864 −0.669 −0.766 −8, 20, −24 L OFC 5.53 143 0.048 Positive 1.527 1.136 0.620 0.817ACC, anterior cingulate cortex; L LP, left lateral parietal cortex; L, left; MNI, Montreal Neurologic Institute; MPFC, medial prefrontal cortex; OFC, orbitofrontal cortex; PCC, posterior cingulate/precuneus cortex; R LP, right lateral parietal cortex; R, right; SFG, superior frontal gyrus; Vent Med FC, ventromedial frontal cortex.
Analyses controlled for age, sex, and WASI-II IQ scores.
Greater connectivity between the DMN and the precuneus (PC) region predicted fewer minutes (R2 = 0.65) and lower percent (R2 = 0.57) of the sleep period spent in stage N1 light sleep (Table 1 and Fig. 1c).
N2Although rsFC of the DMN seed was not significantly correlated with the absolute number of minutes of stage N2 sleep, there were two patterns of connectivity that predicted the percentage of the sleep period in N2. First, a greater percent of time in N2 was predicted by greater positive connectivity between the DMN and right medial prefrontal cortex (R2 = 0.65) but predicted by anticorrelated activity between DMN and the insular cortex (IC; R2 = 0.67; Table 1).
N3DMN whole brain connectivity did not predict subsequent number of minutes or percent of the sleep period spent in stage N3 deep sleep.
Rapid eye movementAs shown in Table 1 and Fig. 1d, positive connectivity between the DMN and a region of the left lateral orbitofrontal cortex (OFC) was predictive of the number of minutes of REM sleep (R2 = 0.77) and the percentage of the sleep period spent in REM (R2 = 0.77).
Total sleep timeGreater TST was predicted by positive connectivity between the DMN and several cortical regions, including the left IC (R2 = 0.65), left OFC (R2 = 0.74), left superior frontal gyrus (SFG; R2 = 0.77), and a region comprising right SFG and anterior cingulate cortex (ACC; R2 = 0.70; Table 1 and Fig. 1e).
Sleep efficiencySleep efficiency was predicted by a combination of anticorrelated connectivity between DMN and brainstem/pons (R2 = 0.80) and positive connectivity with the left OFC (R2 = 0.71; Table 1 and Fig. 1f).
DiscussionConsistent with our hypothesis, we found that rsFC between the DMN and several brain regions was predictive of subsequent overnight sleep as measured by PSG. Specifically, in this correlational study, greater positive connectivity between the DMN and anterior cortical regions (i.e. OFC, SFG, ACC, IC) was associated with greater time spent in more restorative aspects of sleep including greater REM sleep and TST. Similarly, greater connectivity between the DMN and adjacent cortical regions such as the AG and precuneus was predictive of less time spent in non-restorative PSG stages including wake and light N1 sleep. Interestingly, we also found that SE was associated with a combination of greater connectivity between DMN and OFC and anticorrelated connectivity with the brainstem/pons. These findings suggest that greater sleep quality and quantity is predicted by greater congruence of pre-sleep brain activity between regions involved in higher-order cognition and those involved in self-reflective thought, rumination, and worry.
These findings may be best understood in light of the typical effects of increasing homeostatic sleep pressure (e.g. due to sleep deprivation or prolonged wakefulness) on the DMN. Although the DMN is typically most active during resting wake periods, once the period of wakefulness approaches or is extended beyond normal bedtime, the homeostatic pressure for sleep accumulates and there is a corresponding suppression of activity within the DMN [15,16]. The positive (congruent) connectivity we found between DMN and anterior cortical regions implies that as the DMN is suppressed by mounting sleep pressure, these other regions are correspondingly reduced in activation as well. Here, such a pattern was found to predict better sleep later the same night, as measured by greater REM and TST. As the anterior cortical regions tend to be associated with high-level cognitive processing, executive control, and interoceptive awareness [17,18], it follows that congruent reduction of these processes in conjunction with the suppression of self-directed cognitive processes of the DMN (e.g. rumination) would likely facilitate better sleep.
Similarly, greater connectivity between the DMN and adjacent cortical areas (i.e. right AG and the PC) was associated with reduced time in wake or in non-restorative N1 sleep. The AG is known as a cross-modal hub for multisensory integration [19] that has been linked to the experience of consciousness [20]. It therefore follows that reduced AG activation coincident with reduced DMN activation (i.e. functional connectivity) would be associated with reduced conscious wakefulness during the sleep period. Similarly, connectivity between the DMN and a region of the PC that was proximal to the posterior-medial DMN node was associated with less time and percentage of the total sleep period in N1 sleep, the lightest and least restorative sleep stage, which is generally considered to be transitional between wake and deeper stages [21]. The PC has been shown to play an important role in autobiographical memory [22] and integrating information from multiple networks, particularly at rest [23]. It follows then that if the activation of the adjacent PC region is reduced concomitantly with the DMN, this could be associated with reduced activation of autobiographical memory and connectivity with other brain regions that interfere with the ability to fall into deeper sleep. Together, greater connectivity between DMN and these cortical areas tends to be associated with reduced time in wake or the lightest stages of sleep.
Interestingly, SE was predicted by a combined pattern of connectivity with the DMN, including positive connectivity with the left OFC and negative connectivity with the pons. As described above for the role of OFC connectivity in TST, the correlated activity between the DMN and OFC would predict better SE. However, better SE was also simultaneously associated with anticorrelated connectivity to the pons. This suggests that with accumulating sleep pressure, suppression of DMN activation would also be associated with increased activation of the pons, which then predicts greater SE. This is consistent with human research showing that deep brain stimulation of the pons was associated with nearly twice as much REM activity than during the sham condition [24].
It is important to note that we did not find any association between pre-sleep DMN to voxel connectivity and deep N3 sleep. Prior research has shown that within-network connectivity of the DMN is significantly reduced during slow-wave sleep [25]. Moreover, since deep sleep is typically not associated with active cognitions or ruminations, connectivity of the DMN during pre-sleep wakefulness may not be particularly relevant to that stage. However, that speculation will require further research. This study was also limited by our unitary focus on rsFC. Here, we did not utilize any form of cognitive task or comparison group of healthy controls, which might provide additional understanding of the associations between functional connectivity and sleep parameters. Likewise, this study was correlational in nature, so causality cannot be inferred. Future work may consider methods to manipulate these patterns of connectivity to directly examine their influence via experimental methods. Additionally, it is also important to note that the MRI scanner environment is loud, confined, and not inherently restful, which is clearly not a typical situation for most people to undergo during the hours immediately preceding sleep. Further, it is likely that laying for an extended period of time and allowing one’s thoughts to drift in such an unusual situation could induce anxiety, stress, or atypical thoughts that might influence the results. Such possibilities should be considered when interpreting these findings. Future work could include anxiety metrics during the scan to assess such possibilities. Finally, the present findings should be considered in light of the modest sample size and the fact that all participants exceeded clinical scale cutoffs for symptoms typical of insomnia. It is possible that different results would be observed among persons without evidence of insomnia or sleep-related problems.
ConclusionOverall, greater positive connectivity between waking DMN activation and several cortical regions involved in executive functioning, consciousness, and complex cognition was associated with more time in restorative stages of sleep and less time spent awake or in transitional light stages of sleep later the same night. These findings suggest that rsFC of the DMN at bedtime is predictive of objective measures of subsequent sleep, raising the possibility that modulation of the DMN could potentially prove useful for improvement of sleep.
AcknowledgementsThis study was supported by a U.S. Army Medical Acquisition Activity (USAMRAA) Grant (W81XWH-20-1-0173) (WDSK).
Conflicts of interestThere are no conflicts of interest.
References 1. Morin CM, Drake CL, Harvey AG, Krystal AD, Manber R, Riemann D, et al. Insomnia disorder. Nat Rev Dis Primers 2015; 1:15026. 2. Zhou HX, Chen X, Shen YQ, Li L, Chen NX, Zhu ZC, et al. Rumination and the default mode network: meta-analysis of brain imaging studies and implications for depression. Neuroimage 2020; 206:116287. 3. Raichle ME. The brain’s default mode network. Annu Rev Neurosci 2015; 38:433–447. 4. Holub F, Petri R, Schiel J, Feige B, Rutter MK, Tamm S, et al. Associations between insomnia symptoms and functional connectivity in the UK Biobank cohort (n = 29,423). J Sleep Res 2023; 32:e13790. 5. Schiel JE, Holub F, Petri R, Leerssen J, Tamm S, Tahmasian M, et al. Affect and arousal in insomnia: through a lens of neuroimaging studies. Curr Psychiatry Rep 2020; 22:44. 6. Marques DR, Gomes AA, Caetano G, Castelo-Branco M. Insomnia disorder and brain’s default-mode network. Curr Neurol Neurosci Rep 2018; 18:45. 7. Regen W, Kyle SD, Nissen C, Feige B, Baglioni C, Hennig J, et al. Objective sleep disturbances are associated with greater waking resting-state connectivity between the retrosplenial cortex/ hippocampus and various nodes of the default mode network. J Psychiatry Neurosci 2016; 41:295–303. 8. Gagnon C, Belanger L, Ivers H, Morin CM. Validation of the insomnia severity index in primary care. J Am Board Family Med 2013; 26:701–710. 9. Buysse DJ, Reynolds CF 3rd, Monk TH, Berman SR, Kupfer DJ. The Pittsburgh sleep quality index: a new instrument for psychiatric practice and research. Psychiatry Res 1989; 28:193–213. 10. Johns MW. A new method for measuring daytime sleepiness: the Epworth sleepiness scale. Sleep 1991; 14:540–545. 11. Wechsler D. WASI: Wechsler Abbreviated Scale of Intelligence. Second Edition (WASI-II). NCS Pearson; 2011. 12. Rechtschaffen A, Kales A. A manual of standardized terminology, techniques, and scoring systems for sleep stages of human subjects. Brain Information/Brain Research Institute UCLA; 1968. 13. Behzadi Y, Restom K, Liau J, Liu TT. A component based noise correction method (CompCor) for BOLD and perfusion based fMRI. Neuroimage 2007; 37:90–101. 14. Chai XJ, Castanon AN, Ongur D, Whitfield-Gabrieli S. Anticorrelations in resting state networks without global signal regression. Neuroimage 2012; 59:1420–1428. 15. Gujar N, Yoo SS, Hu P, Walker MP. The unrested resting brain: sleep deprivation alters activity within the default-mode network. J Cogn Neurosci 2010; 22:1637–1648. 16. Thomas M, Sing H, Belenky G, Holcomb H, Mayberg H, Dannals R, et al. Neural basis of alertness and cognitive performance impairments during sleepiness. I. Effects of 24 h of sleep deprivation on waking human regional brain activity. J Sleep Res 2000; 9:335–352. 17. Friedman NP, Robbins TW. The role of prefrontal cortex in cognitive control and executive function. Neuropsychopharmacology 2022; 47:72–89. 18. Wang X, Wu Q, Egan L, Gu X, Liu P, Gu H, et al. Anterior insular cortex plays a critical role in interoceptive attention. Elife 2019; 8:e42265. 19. Seghier ML. The angular gyrus: multiple functions and multiple subdivisions. Neuroscientist 2013; 19:43–61. 20. Legostaeva L, Poydasheva A, Iazeva E, Sinitsyn D, Sergeev D, Bakulin I, et al. Stimulation of the angular gyrus improves the level of consciousness. Brain Sci 2019; 9:103. 21. Shrivastava D, Jung S, Saadat M, Sirohi R, Crewson K. How to interpret the results of a sleep study. J Community Hosp Intern Med Perspect 2014; 4:24983. 22. Addis DR, McIntosh AR, Moscovitch M, Crawley AP, McAndrews MP. Characterizing spatial and temporal features of autobiographical memory retrieval networks: a partial least squares approach. Neuroimage 2004; 23:1460–1471. 23. Utevsky AV, Smith DV, Huettel SA. Precuneus is a functional core of the default-mode network. J Neurosci 2014; 34:932–940. 24. Lim AS, Moro E, Lozano AM, Hamani C, Dostrovsky JO, Hutchison WD, et al. Selective enhancement of rapid eye movement sleep by deep brain stimulation of the human pons. Ann Neurol 2009; 66:110–114. 25. Horovitz SG, Braun AR, Carr WS, Picchioni D, Balkin TJ, Fukunaga M, et al. Decoupling of the brain’s default mode network during deep sleep. Proc Natl Acad Sci U S A 2009; 106:11376–11381.
Comments (0)