The hypothesis that ROS and RNS represent keystone mechanisms regulating a variety of physiological processes ranging from neuronal development and structural adjustments to synaptic transmission and neuroplasticity, as well as triggering oxidative injury within the nervous system, is of current and particular interest. The involvement of ROS in physiological conditions and the overall redox balance in the regulation of cytoskeletal modification, neural polarity, signaling modulation, and synaptic transmission has been demonstrated [
95]. The redox equilibrium is also known to be implicated in the physiological processes of neuroplasticity such as neurogenesis, synaptogenesis, and neurochemical changes of the central nervous system (CNS), which express the healthy brain’s ability to adapt its structures and functions in response to environmental changes, or the need to learn new skills through forming new synaptic connections and deconstructing old ones [
96]. Evidence has been provided for the role of ROS/RNS following neuronal activity and in the long-lasting increase in synaptic efficiency, the long-term potentiation (LTP) and long-lasting decrease in the strength of synaptic transmission, and long-term depression (LTD) [
97]. Overall, the neuronal redox equilibrium is designed as a dynamic mechanism which is an integral part of the physiological processes preserving homeostasis and the complex functions of nervous system. Current findings reinforce the results of studies carried out over the last decade and emphasize the role of a controlled generation and release of ROS/RNS, which is operated by all the complex subcellular structures of neurons, in the processes of neuronal remodeling, intracellular signaling, synaptic transmission, and the communication between neurons and glial cells, and open the stage for the development of novel, interventional, protective procedures that many natural compounds might potentially promote by interfering with specific sites of the signaling pathways involved in brain plasticity [
57,
98,
99,
100]. In a damaged brain, as in ischemic stroke, the critical conditions brought about by neuronal degeneration, edema, and inflammation strongly influence the brain’s capacity to restore the lost functions previously performed by the damaged area, and spontaneous recovery proceeds with compensatory plasticity mechanisms based on the reorganization of neural circuits, new functional communications in the remaining neuronal circuits, and enhancing neuronal activity in pre-existing damaged networks [
101]. After an ischemic injury, the NVU loses its function, BBB permeability is enhanced, and multiple mechanisms trigger a cascade of events starting from the decrease in blood flow in the ischemic core until the processes that concur to repair damaged neurons are halted. In the penumbra zone surrounding the ischemic core, the rapid rise in ROS and RNS generation caused by the acute phase of ischemic injury overwhelms antioxidant defenses, resulting in a substantial redox unbalance that is further exacerbated upon reperfusion. The consequent oxidative-nitrosative stress and inflammatory cytokines released by activated microglia and astrocytes critically affect various structural and functional targets, which also predisposes the sufferer to an increased susceptibility of stroke recurrence and other cardiovascular events [
101]. Currently, there is general agreement that certain post-ischemic stroke mechanisms may play an apparently opposite role, which is either beneficial or harmful. ROS and RNS, particularly NO, are usually considered neurotoxic chemical species exerting their detrimental effects via the oxidation of essential macromolecules (DNA, RNA, and proteins) and lipid peroxidation [
44]. Under a steady-state equilibrium between oxidants and antioxidants, as occurs in healthy conditions, ROS and NO may represent suitable mediators for accomplishing the signaling functions required to maintain adequate cell–cell signaling within the NVU [
95,
96,
97]. In ischemic conditions, as occur in ischemic stroke, the oxidative stress due the overwhelming of the ROS and NO associated with lowered antioxidant potential may provide an adequate environment for the regeneration and repair of the damaged NVU, thus mediating parallel processes of neuroplasticity [
102]. The post-ischemic reorganization of cortical representational maps that requires a long-lasting increase in synaptic efficiency (LTP) may explain the apparently controversial role of ROS/RNS, in which the oxidant long-lasting cytotoxic mechanism for both exacerbating neuronal damage (clearance of debris and dead cells) and mediating remodeling cell–cell signaling (neurogenesis and angiogenesis) is essential. It is interesting to note the feedback loop between excessive ROS production and reduced antioxidants availability. ROS generation is somehow allowed by weak antioxidant defense, which is then required for the radicals to burst during the electron transfer step of oxidative metabolism. In this regard, it is intriguing to note that Nrf2 activation is induced by excessive ROS production after a stroke, and that Nrf2 protects the brain against ischemia/reperfusion injury primarily by inducing its target antioxidant genes to counteract excessive ROS production [
103]. Experimental findings have shown that the neuroprotective influence of resveratrol is associated with the activation of the Nrf2 pathway by increasing the expression and activity of superoxide dismutases, glutathione peroxidases, catalase, and reducing lipid peroxidation in brain tissue. Furthermore, the activation of the Nrf2 pathway induced by resveratrol administration inhibits neuroinflammation, apoptosis, oxidative stress, and strokes [
104,
105]. A recent review underlined the possible mechanisms by which peripheral immune components may influence neuronal repair after a stroke, and emphasis is provided on the potential role of metabolites of the gut microbiome in stroke recovery through immunological repair processes, thus outlining an interesting scenario in which malnutrition and dysbiosis can intervene in the processes of plasticity and stroke outcome [
106].

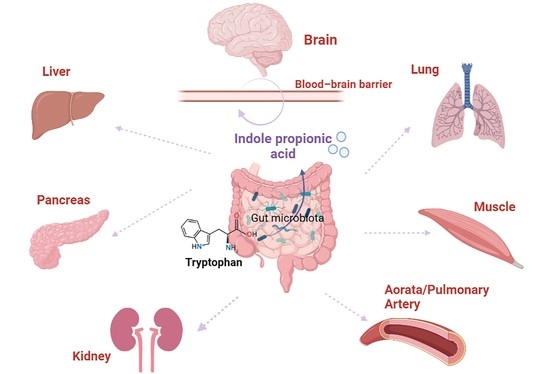
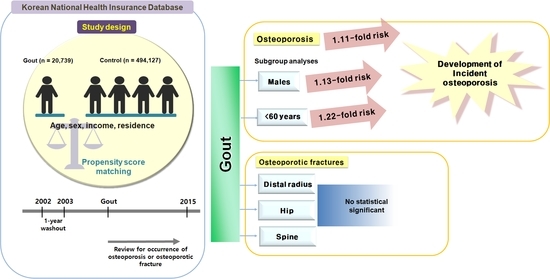
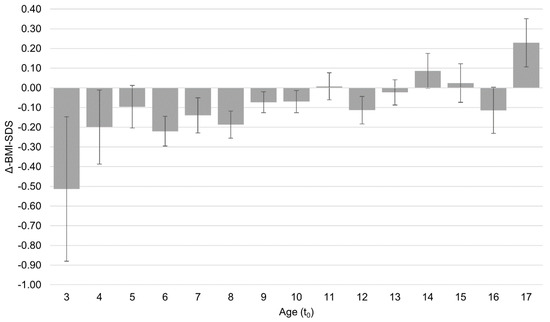
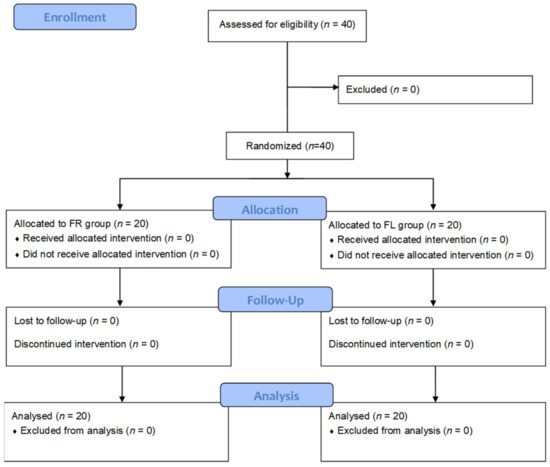


Comments (0)