
Remember me
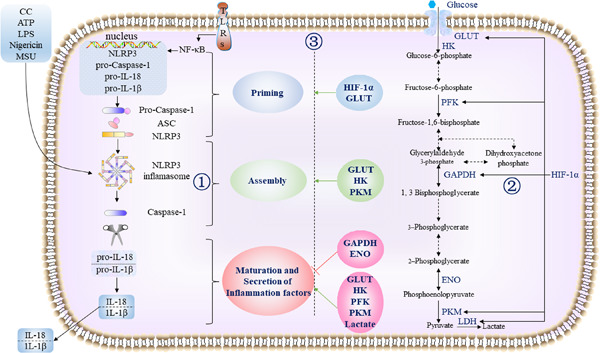
A switch of oxidative phosphorylation to aerobic glycolysis is a possible respond of immune cells to inflammation.42 It has been suggested that macrophages undergo metabolic reprogramming to maintain immunologic and defensive functions and proinflammatory macrophages up-regulate rate of glycolysis rapidly,43 but its role in inflammasome activation is ambiguous.44 It was reported that the canonical activation of NLRP3 inflammasomes depends on glycolysis.45 In contrast, other studies have suggested that the inhibition of glycolytic enzymes such as GAPDH, ENO1, and HK leads to NLRP3 inflammasome activation.20, 46 Meanwhile, NLRP3 inflammasome activation also regulates a series of glycolytic enzymes thereby affecting the corresponding process of glycolysis.
3.1 Glycolysis may regulate the activation of macrophage NLRP3 inflammasomeGlycolysis presents a major regulatory effect in the activation of NLRP3 inflammasome.45, 47 During glucose metabolism reprogramming in macrophages, the switched activation of key glycolytic enzymes is key in regulating NLRP3 inflammasome activation. However, it is still unclear whether glycolytic cascade regulates NLRP3 inflammasome activity positively or negatively.20, 45, 48, 49 Several glycolysis regulators have been involved in NLRP3 inflammasome activation (Figure 1). Previous research has established that some small molecule compounds could regulate glycolysis in macrophages and affect the activation of NLRP3 inflammasome (Table 1).

Schematic representation of the possible mechanism of glycolysis regulates NLRP3 inflammasome in macrophages. ① NLRP3 inflammasome activation is a two-step process, with both priming and activation and NLRP3 must be primed before activation. In priming stage, an NF-κB–activating stimulus, such as LPS binding to TLRs, induces high expression of NLRP3, pro-Caspase-1, pro-IL-1β, and pro-IL-18, which leads to increased expression of their proteins. After priming, canonical NLRP3 inflammasome activation requires a second, distinct signal to activate NLRP3 and lead to the formation of the NLRP3 inflammasome complex. NLRP3 specific ligands can also activate NLRP3 inflammasome. As a result, pro-caspase-1 is converted to Caspase-1. Upon activation, active Caspase-1 cleaves the pro-IL-1β and pro-IL-18 into their mature forms, which secret out of cells. ② Glycolysis is a biological process that occurs to convert glucose into pyruvate to provide energy for cells. Since the glycolysis cycle involves the conversion of blood sugar into an anion of pyruvic acid (pyruvate), glycolysis is also referred to as the citric acid cycle under hypoxia condition or aerobic glycolysis. Aerobic glycolysis refers to the process of glycolysis under aerobic conditions. When aerobic glycolysis occurs under hypoxic conditions, it called anaerobic glycolysis. There are ten reactive steps to occur that involve several catalyst enzymes such as HK, PFKKM and PKM and intermediate compounds. HIF-1a is a key regulator of glycolysis during hypoxia, upregulate the coding of aerobic glycolysis enzyme at the transcription level in macrophages. ③ HIF-1a, GLUT, HK, and PK induce the priming step of NLRP3 inflammasome. GLUT, HK, and PK promote NLRP3 inflammasome assembly. GLUT, HK, PFK, PK, and lactate, promote inflammatory factors secretion mediated by NLRP3 inflammasome activation. GAPDH and ENO inhibit inflammatory factors secretion mediated by NLRP3 inflammasome activation. ASC, apoptosis-associated speck-like protein; ENO, enolase; GAPDH, glyceraldehyde-3-phosphate dehydrogenase; GLUT, glucose transporter;, HIF-1a, hypoxia inducible factor-1α; HK, hexokinase; IL-18, interleukine-18; IL-1β, interleukine-1β; LDH, lactate dehydrogenase; LPS, lipopolysaccharide; NF-κB, nuclear factor κβ; NLRP3, Nod-like receptor protein 3; PFK, phosphofructokinase; PKM, pyruvate kinase muscle isozyme; TLRs, Toll-like receptors
Table 1. Some compounds regulate glycolysis and then inhibit NLRP3 inflammasome-dependent inflammation in macrophages Target Compound Mechanism Effect on NLRP3 References HK 2-DG Promotes HK2 dissociation from the outer mitochondrial membrane Inhibits the expression of caspase-1, IL-1β and IL-18 [12] ATRA Enhances HK2 expression Reduces NLRP3 inflammasome-dependent IL-1β secretion [12] Andrographolide Inhibits the activity of HK2 Reduces the release of IL-1β [8] PK Shikonin Inhibits EIF2AK2 phosphorylation Reduces caspase-1 activity, and IL-1β and IL-18 release [12] LBP Reduces the expression of PKM2 protein Reduces IL-1β production [10] DET Inhibits the nuclear localization of PKM2 Attenuates the release of IL-1β [14] IRD Targets PKM2 and inhibits its downstream expression Inhibits the release of IL-1β [54] HIF-1α Chaetocin Inhibits HIF-1α expression Suppresses priming of NLRP3 inflammasome and IL-1β synthesis [55] GAPDHGB111-NH2
KB
Inhibits GAPDH expression, thereby decreases glycolytic flux Inhibits inflammatory factors maturation and release [20] α-enolaseGB111-NH2
EB
Inhibits α-enolase expression, then decreases glycolytic flux Induces inflammatory factors maturation and release [9] LDHA GSK2837808a Effective and selective inhibitor of lactate dehydrogenase A (LDHA) Reduces the protein levels of mature IL-1β and active caspase-1 [11] Abbreviations: DET, deoxyelephantopin; GAPDH, glyceraldehyde-3-phosphate dehydrogenase; HIF-1a, hypoxia inducible factor-1α; HK, hexokinase; IL-18, interleukine-18; IL-1β, interleukine-1β; LBP, Lycium barbarum polysaccharide; NLRP3, Nod-like receptor protein 3; PK, pyruvate kinase; PKM, pyruvate kinase muscle isozyme. 3.1.1 GLUTAs the most abundant glucose transporter in macrophages,31 GLUT1 induces the full activation of NLRP3 inflammasome. Its silence by pharmacological inhibition (STF-31: iGLUT1) or gene knockdown results in attenuated phorbol myristate acetate (PMA)-induced gene expressions of both NLRP3 and IL-1β, suggesting that GLUT1 involves in the priming step of NLRP3 inflammasome.32 Additionally, inhibition of GLUT1 reduces glucose uptake and restrains glycolysis in macrophages, prevents ASC speck formation and NLRP3 inflammasome assembly.32 GLUT1 knockdown also reduces maturation and release of IL-1β in macrophages.56 The reduction of GLUT1 in vivo is regulated by the proto-oncogene Casitas B-lineage lymphoma (Cbl) through posttranscriptional modification. Cbl encodes an ubiquitin ligase, and mediates protein ubiquitination.57 The expression of GLUT1 is upregulated, thereby promotes maturation and secretion of IL-1β and actives caspase-1 in THP-1-derived macrophages with Cbl-KO. In summary, the above research shows that GLUT1 helps in both of the priming and activation of the NLRP3 inflammasome.
3.1.2 HKHK, as the first speed-limiting enzyme in the glycolytic pathway, activates NLRP3 inflammasome to promote the maturation and secretion of inflammatory factors. So far, HK isozymes, named from HK1 to HK4, have been identified in mammals.58 HK can be incorporated into the outer membrane of mitochondria through the interaction with the voltage-dependent anion channel (VDAC) at, a pattern recognition receptor.46 Among the three VDAC identified subtypes (VDAC1-3), VDAC1 is widely expressed in a vast many cell types.59 Dissociation of HK from VDAC1 induces inflammasome assembly, then the maturation and release of IL-1β and IL-18. HK1 knockdown inhibits caspase-1 activation, IL-1β and IL-18 release stimulated by LPS/ATP in murine macrophages J774A.1.45 Decreased expression of HK1 mitigates caspase-1 activation, therefore reducing secretion of IL-1β and IL-18 in wild type bone marrow–derived macrophages (BMDMs) in response to joint stimulation by LPS and ATP.45 Some agents that inhibit HK dissociating from the mitochondrial membrane or expression suppress NLRP3 inflammasome activation. As a derivative of vitamin A, all-trans retinoic acid (ATRA) enhances HK2 expression, and converts the cellular metabolism of macrophages to glycolysis in LPS treatment, which promotes NLRP3 inflammasome activation. What's more, 3-bromopyruvate (3BP), as selective inhibitor of HK2, attenuates IL-1β secretion induced by ATRA in macrophages.50 As the major negative regulator of autophagy,60, 61 mammalian target of rapamycin complex 1 (mTORC1) induces HK1 expression via phosphorylation of 4E-binding protein 1(4E-BP1), the eukaryotic translation initiation factor.45, 60, 62 In line with this, HK1 expression can be inhibited by Torin1, the mTOR selective inhibitor63 or rapamycin64 or knockdown the 4E-BP1 and Raptor, a scaffold for mTORC1 complex substrates.65 Carbon monoxide-releasing molecules (CORM) inhibits mTORC1 activation, thereby inhibits glycolysis during NLRP3 inflammasome activation,66 inhibits ASC oligomerization and caspase-1 activation, and decreases secretions of IL-1β and IL-18 in macrophages stimulated by LPS and ATP. Elevated levels of glucose-6-phosophate (G6P) also promotes HK release from mitochondria, thus slows the rate of G6P production.67, 68 Acting as a d-glucose mimic, 2-deoxyglucose (2-DG) is phosphorylated by HK to 2-deoxyglucose-6-phosphate (2-DG6P), which inhibits the function of HK in a way similar to G6P. As an immune checkpoint molecule, T cell immunoglobulin and mucin domain-3 (Tim-3) inhibits mRNA and protein expression of HK2 in RAW264.7 macrophages via the STAT1 pathway, thereby inhibiting the mRNA level of IL-1β and protein level of pro-IL-1β. When knocked down the HK2 expression using HK2 siRNA, the inhibition is reversed.69 Synthesis of novel andrographolide Beckmann rearrangement derivatives can inhibit the activity of HK2, thereby reducing the release of IL-1β in RAW264.7 cells induced by LPS.51 In summary, HK contributes to NLRP3 inflammasome priming and activation; the agents that inhibit HK expression or dissociating from the mitochondrial membrane may prevent NLRP3 inflammasome-mediated inflammation.
3.1.3 PFKPFK1 is one of the rate-limiting enzymes in glycolysis, which can convert fructose 6-phosphate into fructose 1,6-phosphate.70 PFK1 has three isoforms: platelet (PFKP), muscle (PFKM), and liver (PFKL).70 PFKM is predominantly expressed in normal muscle and neuronal tissues.71 The long 3'-UTR on the mRNA of PFKM can bind to microRNA-21 and restrict the expression of PFKM, thus affecting the secretion of IL-1β in BMDMs stimulated by Mycobacterium tuberculosis (Mtb).72 PFKFB3 synthesizes fructose 2, 6-bisphosphate, which act as a powerful allosteric activator of PFK1.73 Transcription of PFKFB3 is enhanced by binding of the PFKFB3 promoter to Zinc fingers and homeoboxes (Zhx2), thereby promoting the release of IL-1β in LPS-stimulated BMDMs.74 Microglia are macrophages that reside in the central nervous system.75 Monocarboxylate transporters (MCTs) are also important glycolytic regulators that transport excess lactate out of cells. PFKFB3 can be promoted by MCT1 through Hif-1α, thus promoting the secretion of IL-1β by LPS-stimulated primary and BV2 microglia. Overexpression of PFKFB3 rescued the siMCT1-mediated reduction of IL-1β expression and demonstrated the promoting effect of PFKFB3 on the release of IL-1β from LPS-stimulated microglia.76 In summary, the expression of PFK1 mainly promotes the maturation and secretion of inflammatory factors.
3.1.4 PKPK is the key enzyme in the last step of glycolysis, which catalyzes the reaction between phosphoenolpyruvate (PEP) and ADP to form pyruvate and ATP. PKM2 regulates the transcription of GLUT1, LDHA and other glycolysis-related genes and ties to hypoxia inducible factor-1α(HIF-1α)to promote aerobic glycolysis in LPS-treated macrophages.77 PKM2 induces the assembly of NLRP3 inflammasomes and release of inflammatory factors by phosphorylation of factor 2 alpha kinase 2 (EIF2AK2), which is well known to play a critical role in inflammasome activation.47, 78, 79 PKM2 knockdown or its inhibitor shikonin suppresses EIF2AK2 phosphorylation,80 thereby reducing casapase-1 activity, inhibiting release of IL-1β or IL-18 in BMDMs stimulated by LPS and the NLRP3 inflammasome activator ATP.47 And interaction between NLRP3 and ASC (also termed as PYD and CARD domain containing PYCARD) is inhibited upon PKM2 knockdown in caspase-1−/− BMDMs induced by LPS and ATP.47 It indicates that PKM2 regulates the production and release of proinflammatory factors in the NLRP3 inflammasome-dependent pathway through phosphorylation of EIF2AK2. As the product of glycolysis, lactate contributes to the inflammatory process through multiple mechanisms.51, 70 For example, it activates NLRP3 inflammasome-dependent inflammatory release, IL-1β release and EIF2AK2 phosphorylation in BMDMs triggered by LPS.47 EIF2AK2 knockdown or addition of its inhibitor C1681 inhibits the phosphorylation of EIF2AK2 induced by lactate and the release of IL-1β in LPS-activated BMDMs.47 As the main bioactive component of Chinese wolfberry, Lycium barbarum polysaccharide (LBP) can reduce the expression of PKM2 protein in LPS-induced RAW264.7 macrophages, but has no effect on the expression of PKM2 mRNA. LPS inhibited the ubiquitination of PKM2, possibly by downregulating the expression of ubiquitin ligases, including Nedd4L, Nedd4 and Gnb2. LBP interferes with the inhibition of PKM2 ubiquitination by upregulating the expression of Nedd4L, Nedd4, and Gnb2, thereby reducing IL-1β production in LPS-induced RAW264.7 macrophage.52 Deoxyelephantopin (DET), a naturally occurring sesquiterpene lactone from Elephantopus scaber, inhibited the nuclear localization of PKM2 and thus attenuated the release of IL-1β from LPS-stimulated RAW264.7 macrophages.53 IRD is a main isoflavone derived from the root of the plant Belamcanda chinensis (L.) Redouté, can target PKM2 and inhibit its downstream expression of p-JAK1, p-STAT1, p-STAT3, p-p65, iNOS and COX2, thereby inhibiting the release of IL-1β from LPS-treated RAW264.7 macrophages. PKM2 agonist DASA-58 could abolish this inhibitory effect.82 In summary, inhibition of PKM2 expression mainly reduces the maturation and secretion of inflammatory factors.
3.1.5 LactateLactate is the end product of glycolysis and is derived from pyruvate by the enzyme lactate dehydrogenase (LDH).54 Lactate can induce phosphorylation of PKR,47 promoting protein expression of PKR, induces the NLRP3 inflammasome-dependent IL-1β secretion in nigericin, ATP, monosodium urate (MSU) crystals, or alum stimulated BMDM and THP-1 cell.24 The protein levels of mature IL-1β and active caspase-1 were reduced by using 2-DG or LDH inhibitor GSK2837808a, or by lactate dehydrogenase a (LDHA) specific small interfering RNA (siRNA). But the levels of NLRP3, ASC, pro-caspase-1 and pro-IL-1β did not change.24 However, in other studies, increased lactate did not promote the activation of NLRP3 inflammasome. In macrophages and monocytes, exogenous lactate reduced TLR4-mediated induction of IL-1β, NLRP3, and pro-caspase-1; activation of nuclear factor κβ (NF-κB); release of IL-1β; and cleavage of caspase-1.83 Addition of lactate directly inhibits pro-IL-1β, NLRP3, and Caspase-1 levels in LPS-mediated human peripheral blood mononuclear cells, and also inhibited pro-caspase-1 cleavage, mature caspase-1 proteolytic activity, as well as caspase1-dependent process and extracellular release of IL-1β. It is suggested that lactate has no promoting effect on the activation of NLRP3, and has moderate antagonistic effect. Other studies have found that exogenous lactate stimulation can inhibit the expression of PFKFB3 induced by LPS in BV2 cells, and also show inhibitory effect on the expression of IL-1β.76 The results of these experiments are different, the possible difference is in the study of lactic acid, exogenous lactate may act as an exogenous inhibitor of NLRP3, and the increase of lactic acid caused by intervention methods has a promoting effect on the activation of NLRP3 inflammasome.
3.1.6 HIF-1ΑHIF-1α is a decisive mediator of glycolysis, by inducting enzymes in glycolysis, especially HK2, PKM2, glucose-6-phosphate isomerase (GPI) and triosephosphate isomerase.84 HIF-1α promotes priming of NLRP3 inflammasome by upregulating the expression of NLRP3,85 and is important for IL-1β release. Overexpression of HIF-1α and IL-1β are found in THP-1-derived macrophages stimulated by palmitic acid; furthermore, knockdown of HIF-1α can inhibit the proinflammatory effects of palmitic acid. NLRP3 inflammasome is activated and HIF-1α overexpressed in RAW246.7 macrophages stimulated with LPS/ATP; GN44028, as HIF-1α inhibitor, suppresses the expression level of HIF-1α and NLRP3, thus IL-1β release.86 Chaetocin, an antibiotic with epipolythiodioxopiperazines structure produced by Chaetomium sp,87 reduces the level of pro-IL-1β by inhibiting HIF-1α, thereby affecting NLRP3 priming inflammasome and IL-1β synthesis.55
GAPDH and ENO affects glycolytic flux, including glycolytic capacity and glycolytic reserve, which has been considered to involve in the full activation of NLRP3 inflammasome.39, 41 GB111-NH2,88, 89 as a peptide-based compound that inhibits both GAPDH and α-enolase,20 restores metabolism downstream of glycolytic disruption, which is sufficient to suppress the inflammasome response by reinstating NADH generation and reducing mitochondrial ROS generation.20 Furthermore, KA and EB, as inhibitors of GAPDH and α-enolase respectively, also affect glycolytic flux, and then the activation of NLRP3 inflammasome.20 However, inhibition activity of GAPDH or other enzyme in lower glycolysis could disrupt glycolytic flux and induce the activation of the NLRP3 inflammasome in primed murine bone marrow–derived macrophages.20 Taken together, the regulatory roles of glycolytic flux in NLRP3 inflammasome activation are still confused.
3.2 NLRP3 inflammasome activation promotes regulates glycolysis in macrophagesKey enzymes of glycolysis and end product (lactate) from the glycolytic pathway, promote NLRP3 inflammasome activation as mentioned above. Likewise, evidence shows that activation of NLRP3 also regulates glycolysis via several mechanisms.15
As one of the HK isoforms, HK1 expression is induced by NLRP3 inflammasome activator ATP and LPS but not with the AIM2 inflammasome activator poly (dA:dT) in BMDM. It indicates the specificity of NLRP3 inflammasome in activating HK1.45 Caspase-1 cleavage assay in vitro combined with the proteomic analysis of caspase-1 reveals those glycolytic enzymes including aldolase, triose-phosphate isomerase, GAPDH, α-enolase and PK are caspase-1 substrates.19 Expression of GAPDH, aldolase, enolase and TIM was suppressed in peritoneal macrophages from wild-type mice infected with Salmonella typhimurium, but not in macrophages from mice lacking caspase-1.90 The targeting of caspase-1 to glycolytic enzymes was also demonstrated in the diaphragm of mice with LPS-induced septic shock.19 However, these results await further animal experimental confirmation. PFKFB3 synthesizes fructose 2, 6-bisphosphate, which act as a powerful allosteric activator of PFK1,73 thus drive glycolysis.15 In macrophages, LPS and Aβ increased the rate of glycolysis and PFKFB3 expression, and these effects were counteracted by MCC950, a selective NLRP3 inhibitor.91 IL-1β induced glycolysis activation by activation of PFKFB3 mimicking the role of LPS plus Aβ, which added the evidence to support the impact of inflammation on metabolomic profiles. This view was further supported by other observations that PFKFB3 and changes in glycolysis stimulated by LPS + Aβ were attenuated in BMDMs from NLRP3−/− and interleukin-1-receptor type 1-homozygous knockout (IL-1R1−/−) mice.91 In line with this notion, 3PO, the PFKFB3 inhibitor, attenuates glycolysis induced by LPS + Aβ. In summary, the NLRP3 inflammasome could modulate glycolysis by upregulating PFKFB3 expression in an IL-1β-dependent manner in macrophages.15
Research on the regulation of NLRP3 by glycolysis was introduced in-depth above (Figure 2), but the detailed mechanism how NLRP3 regulates glycolysis is still to be explored. Additionally, activation of pro-Caspase-1 induced by cigarette smoke extract results in decrease in basal glycolytic flux and damaged glycolytic burst after LPS stimulation in vitro,92 which implies that the regulatory roles of NLRP3 inflammasome activation in glycolytic flux are controversial.

Schematic representation of the possible mechanism of NLRP3 inflammasome regulates glycolysis in macrophages. Caspase-1 can promote the expression of key glycolytic enzymes HK and PFK and inhibit the expression of ALD, TIM, GAPDH and ENO. IL-1β can promote the activity of PFK. ALD, aldolase; ASC, apoptosis-associated speck-like protein; ENO, enolase; GAPDH, glyceraldehyde-3-phosphate dehydrogenase; GLUT, glucose transporter; HK, hexokinase; IL-18, interleukine-18; IL-1β, interleukine-1β; NLRP3, Nod-like receptor protein 3; PFK, phosphofructokinase; TIM, Triosephosphate isomerase
Comments (0)