Experiment 1
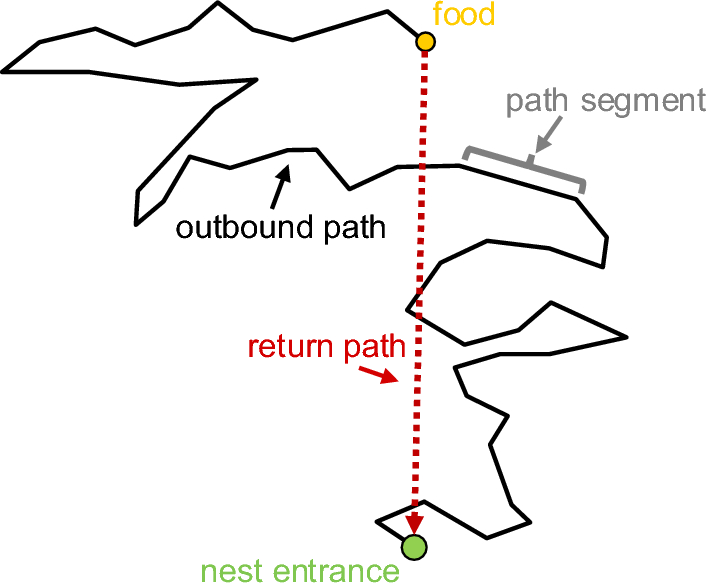
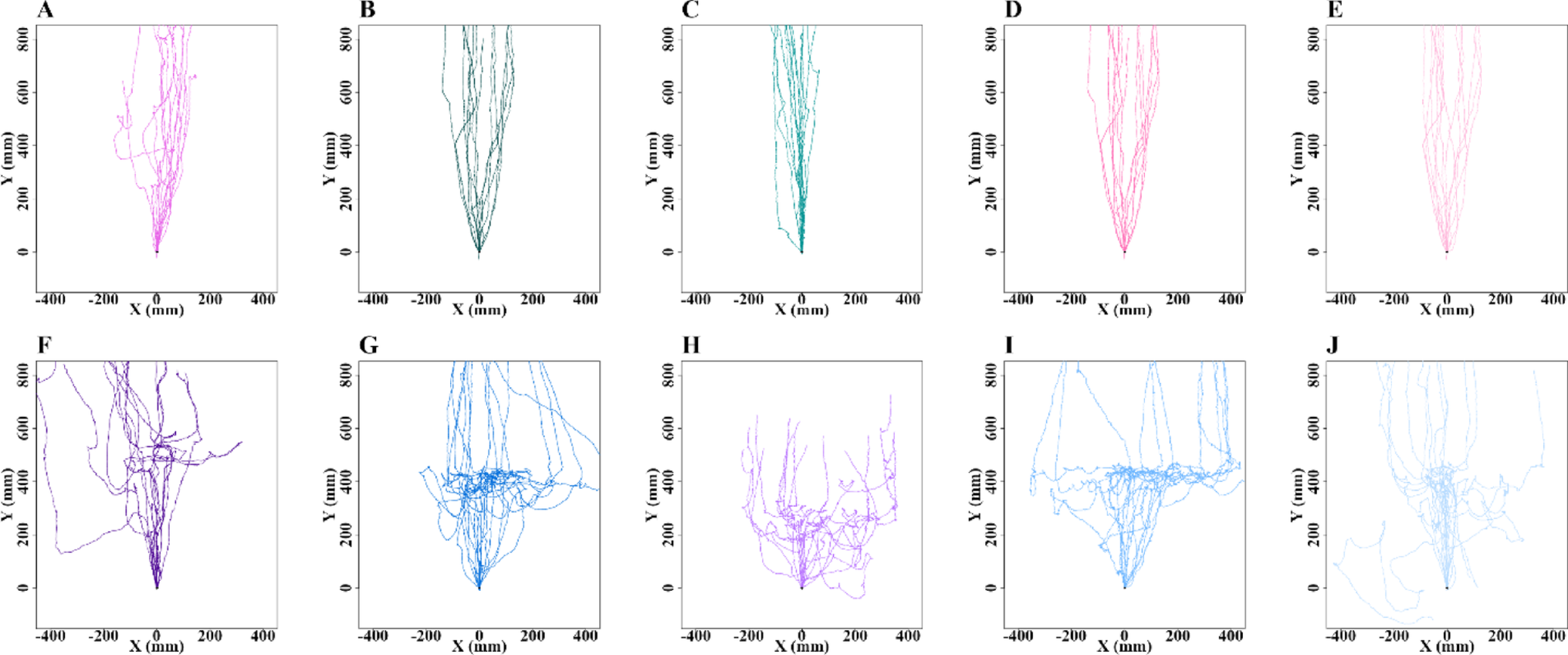
After emerging from the nest during the evening twilight, all the nest-tree foragers followed a stereotypical vertical path up the eucalyptus tree trunk and continued along the same foraging corridor every day. We conducted observations on these experienced foragers for two consecutive nights on their nest trees for control (N = 15) and odour-test (N = 15) purposes. The presence of odours on their foraging corridor resulted in detours for foraging (Fig. 1). Specifically, with Eucalyptus oil, 46.4% of ants walked over the odour, while 53.6% took detours to avoid it. With Olive oil and Flax-seed oil (sources of oleic acid and linoleic acid, respectively), 35% and 40% of foragers, respectively, walked over the odour, while 65% and 60% took detours. With Tea tree oil and Lavender oil, none of the foragers walked over the odour, with 26% and 22% of foragers, respectively, taking a U-turn and returning to the nest, while 74% and 78% took detours away from the odour line. These results suggest that foragers often exhibit a phobic response, with nearly half of them avoiding the familiar Eucalyptus oil, more avoiding oils containing oleic and linoleic acid, and complete avoidance of Tea tree and Lavender oils.
In path characteristics, ants exhibited a more curved and meandering trajectory in the odour-change conditions compared to the control conditions. We found differences across conditions in each of our two measures of path meander. Firstly, sinuosity increased significantly in the odour-change conditions (Olive oil control vs. Olive oil test: t = − 7.4, df = 17.4, P ≤ 0.0001; Flax-seed oil control vs. Flax-seed oil test: t = − 14.1, df = 19.4, P ≤ 0.0001; Tea tree oil control vs. Tea tree oil test: t = − 10.4, df = 18.2, P ≤ 0.0001; Lavender oil control vs. Lavender oil test: t = − 12.2, df = 22.8, P ≤ 0.00001) except for Eucalyptus oil (Eucalyptus oil control vs. Eucalyptus oil test: t = 0.4, df = 23.8, P = 0.6) (Fig. 2A). Secondly, E amax was lower in the odour-change conditions, with the ants having a smaller amount of displacement per unit length travelled compared to the control conditions. The t-tests showed a significant difference in test conditions compared to the control conditions in all comparisons except Eucalyptus oil (Olive oil control vs. Olive oil test: t = 6.1, df = 25.2, P ≤ 0.0001; Flax-seed oil control vs. Flax-seed oil test: t = 7.2, df = 16.8, P ≤ 0.0001; Tea tree oil control vs. Tea tree oil test: t = 9.08, df = 16.9, P ≤ 0.0001; Lavender oil control vs. Lavender oil test: t = 9.6, df = 23.2, P ≤ 0.0001; Eucalyptus oil control vs. Eucalyptus oil test: t = − 0.02, df = 27.5, P = 0.9) (Fig. 2B).
Odour changes had a noticeable impact on the speed, orientation angular velocity, and duration of the foragers. When the foragers sensed non-visual odour stimuli, they slowed down, frequently shifted their gaze in different directions, and stayed in the recording area longer compared to the respective controls (Fig. 3). In speed, the t-tests revealed significant differences in all comparisons (Fig. 3A), with the ants speeding up with Eucalyptus oil and slowing down with other odours (Eucalyptus oil control vs. Eucalyptus oil test: t = − 3.04, df = 27.6, P < 0.01; Olive oil control vs. Olive oil test: t = 6.5, df = 26.6, P ≤ 0.0001; Flax-seed oil control vs. Flax-seed oil test: t = 5.2, df = 27.01, P ≤ 0.0001; Tea tree oil control vs. Tea tree oil test: t = 8.5, df = 24.2, P ≤ 0.0001; Lavender oil control vs. Lavender oil test: t = 13.9, df = 19.06, P ≤ 0.00001). Changes in odour also increased the magnitude of orientation angular velocity in the foragers (Fig. 3B). The t-tests showed significant differences between all pairs in orientation angular velocity except for Olive oil (Eucalyptus oil control vs. Eucalyptus oil test: t = − 4.6, df = 15.2, P ≤ 0.001; Flax-seed oil control vs. Flax-seed oil test: t = − 20.8, df = 15.1, P ≤ 0.00001; Tea tree oil control vs. Tea tree oil test: t = − 8.9, df = 21.1, P ≤ 0.0001; Lavender oil control vs. Lavender oil test: t = − 9.06, df = 25.1, P ≤ 0.0001; Olive oil control vs. Olive oil test: t = 0.3, df = 20.9, P = 0.7). The amount of time spent in the recording area also increased, as foragers stopped just before the odour line and meandered around it (Fig. 3C). Consequently, trips took significantly more time with all odours except for Eucalyptus oil (Olive oil control vs. Olive oil test: t = − 5.3, df = 14.3, P ≤ 0.0001; Flax-seed oil control vs. Flax-seed oil test: t = − 4.8, df = 16.4, P ≤ 0.0001; Tea tree oil control vs. Tea tree oil test: t = − 6.6, df = 14.6, P ≤ 0.0001; Lavender oil control vs. Lavender oil test: t = − 5.1, df = 14.1, P < 0.0001; Eucalyptus oil control vs. Eucalyptus oil test: t = − 0.7, df = 23.4, P = 0.4).
Odour change increased scanning in foragers. In Control recordings, i.e. those without any odour change, the majority of the foragers performed a single scan across their entire recording area (Fig. 4A). In contrast, on odour trials, where ants experienced a non-visual odour change, foragers scanned at least twice, with a maximum of 28 (Fig. 4A). The t-tests revealed significant differences in the number of scanning bouts in all comparisons (Eucalyptus oil control vs. Eucalyptus oil test: t = − 4.7, df = 18.3, P ≤ 0.001; Olive oil control vs. Olive oil test: t = − 4.8, df = 15.1, P ≤ 0.0001; Flax-seed oil control vs. Flax-seed oil test: t = − 4.1, df = 15.9, P ≤ 0.0001; Tea tree oil control vs. Tea tree oil test: t = − 5.5, df = 14.9, P ≤ 0.0001; Lavender oil control vs. Lavender oil test: t = − 4.7, df = 15.9, P ≤ 0.0001). The duration of scanning bouts also increased significantly with odour changes (Fig. 4B), in all comparisons except for Eucalyptus oil (Olive oil control vs. Olive oil test: t = − 4.2, df = 14.4, P ≤ 0.001; Flax-seed oil control vs. Flax-seed oil test: t = − 2.7, df = 27.87, P < 0.01; Tea tree oil control vs. Tea tree oil test: t = − 4.8, df = 14.5, P < 0.001; Lavender oil control vs. Lavender oil test: t = − 5.4, df = 24.1, P ≤ 0.0001; Eucalyptus oil control vs. Eucalyptus oil test: t = − 1.1, df = 17.60, P = 0.2).
Experiment 2
To understand whether the detouring mechanism is triggered by neophobia or inherent aversion to an odour, we applied Lavender oil, which all the ants avoided on the Test day in Experiment 1, and recorded individual ant behaviour over three days. On the control day (N = 16), all ants went straight up the tree. On the first test day (Test-Day1) (N = 16), after applying the lavender oil odour, only one ant walked over the odour, one took a U-turn, and the rest detoured to avoid the odour. On the second test day (Test-Day2) (N = 16), five ants walked over the odour. By the third test day (Test-Day3) (N = 16), only two ants detoured, while the rest walked over the odour (Fig. 5).
To understand the impact of the odour on ants’ navigational performance, we compared the path characteristics of foragers on the Control and Test-Day1 to 3. Firstly, the sinuosity was observed to be low in the Control condition. It increased significantly on Test-Day1, decreased on Test-Day2, and showed variation in magnitude on Test-Day3. The repeated-measures ANOVA model revealed a significant difference (F3,56 = 4.4, P < 0.006) across the conditions (Fig. 6). A priori Helmert Contrast-1 (Z3,56 = 6.1, P = 0.0009) showed significant differences between the control and test conditions. Contrast-2 revealed a trend (Z3,56 = 1.7, P = 0.07), while contrast-3 revealed significant differences between the Test-Day2 and Day3 (Contrast-3: Z3,56 = 3.26, P = 0.002). These results show significant recovery by Test-Day3.
Secondly, E amax as observed to be lower in the Test-Day1 and it gradually increased over the next two Test Days. The repeated-measures ANOVA revealed a significant difference (F3,56 = 7.8, P ≤ 0.0001) across the days (Fig. 6). The first two a priori Helmert tests (Contrast-1: Z3,56 = 7.1, P ≤ 0.0001; Contrast-2: Z3,56 = 5.6, P ≤ 0.0008) showed significant differences between the contrasted conditions, while Contrast-3 revealed no significant differences between the Test-Day2 and Day3 (Contrast-3: Z3,56 = 1.85, P = 0.06). These results indicate that the experimental odour applied on their foraging route had a significant impact on the characteristics of the ants’ paths and that the ants recovered in subsequent trips.
The odour change had a noticeable impact on the speed, orientation angular velocity, and duration of the foragers from the Control Day to Test-Day3. The mean speed of foragers decreased on Test-Day1 and gradually increased over the next two Test Days. The repeated-measures ANOVA revealed a significant difference (F3,56 = 3.8, P < 0.03) across the conditions (Fig. 7). A priori Helmert Contrast-1 (Z3,56 = 4.4, P ≤ 0.0006) showed significant differences between the control and test conditions. Contrast-2 revealed a borderline effect (Z3,56 = 2.2, P = 0.04), while the other Helmert contrast revealed no significant differences between Test-Day2 and Test-Day3 (Contrast-3: Z3,56 = 1.7, P = 0.08).
The orientation angular velocity of foragers increased on Test-Day1 and gradually decreased over the next two Test Days (Fig. 7). The repeated-measures ANOVA revealed a significant difference (F3,56 = 10.2, P ≤ 0.00001) across the days. A priori Helmert test Contrast-1 (Z3,56 = 3.1, P = 0.003), Contrast-2 (Z3,56 = 6.2, P ≤ 0.00006), and Contrast-3 (Z3,56 = 3.03, P ≤ 0.003) all showed significant differences between contrasted conditions. The duration of time spent in the recording area also increased, as foragers stopped just before the odour line and meandered around it. However, the repeated measures ANOVA revealed no significant differences (F3, 56 = 2.1, P = 0.1) across the days (Fig. 7). Overall, these results indicate that the odour treatment significantly influenced the foragers’ speed and orientation angular velocity.






Comments (0)