
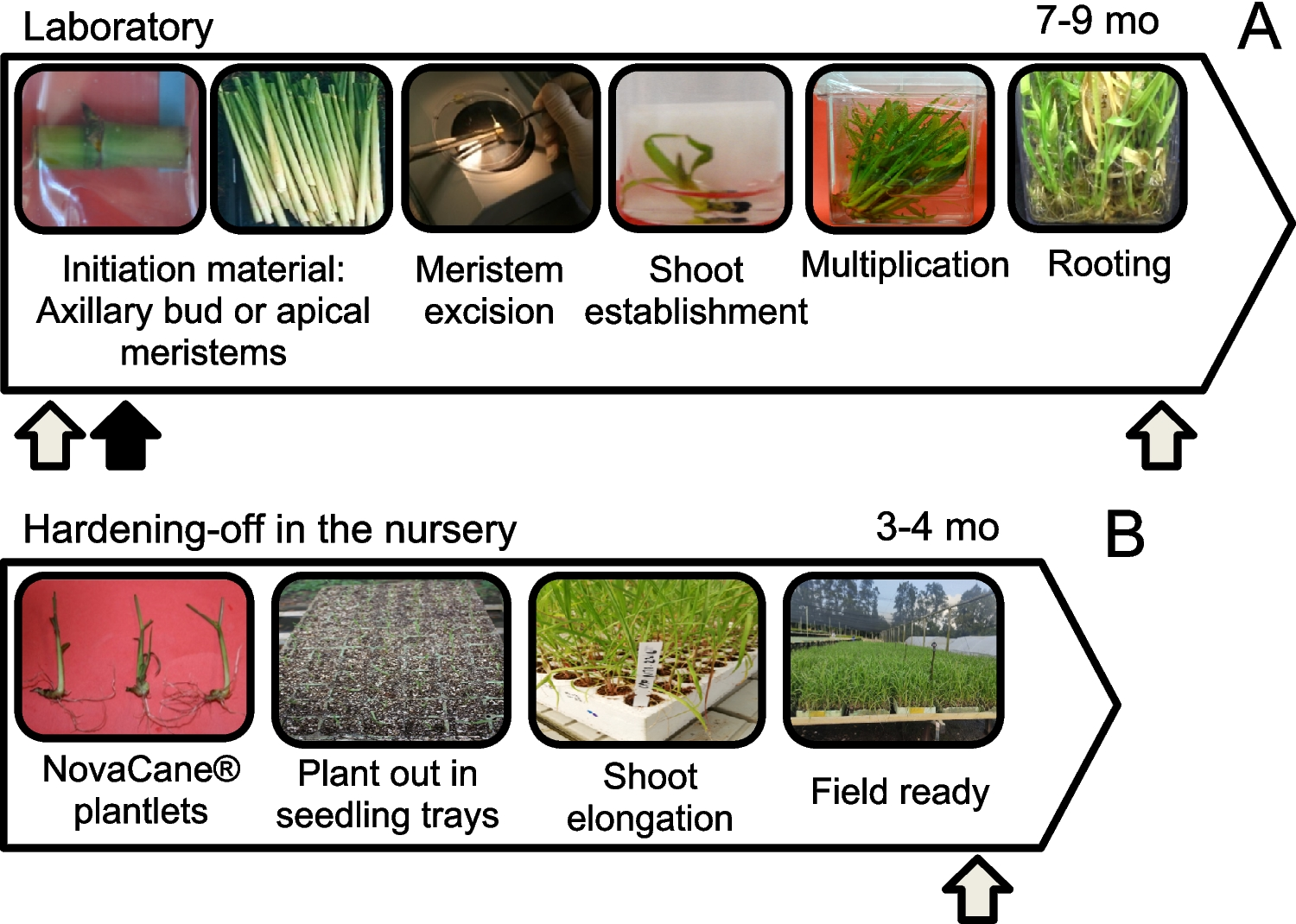
Remember me








B. nigra Diyarbakır ecotype tissue culture studies were carried out in the first phase of the study for the in vitro propagation of the plant. In the study's second phase, different concentrations of Cu were applied to the propagated plants to validate their post-propagation heavy metal accumulation capacity. The Cu content in the roots, stems, and leaves of the plants was determined by AAS.
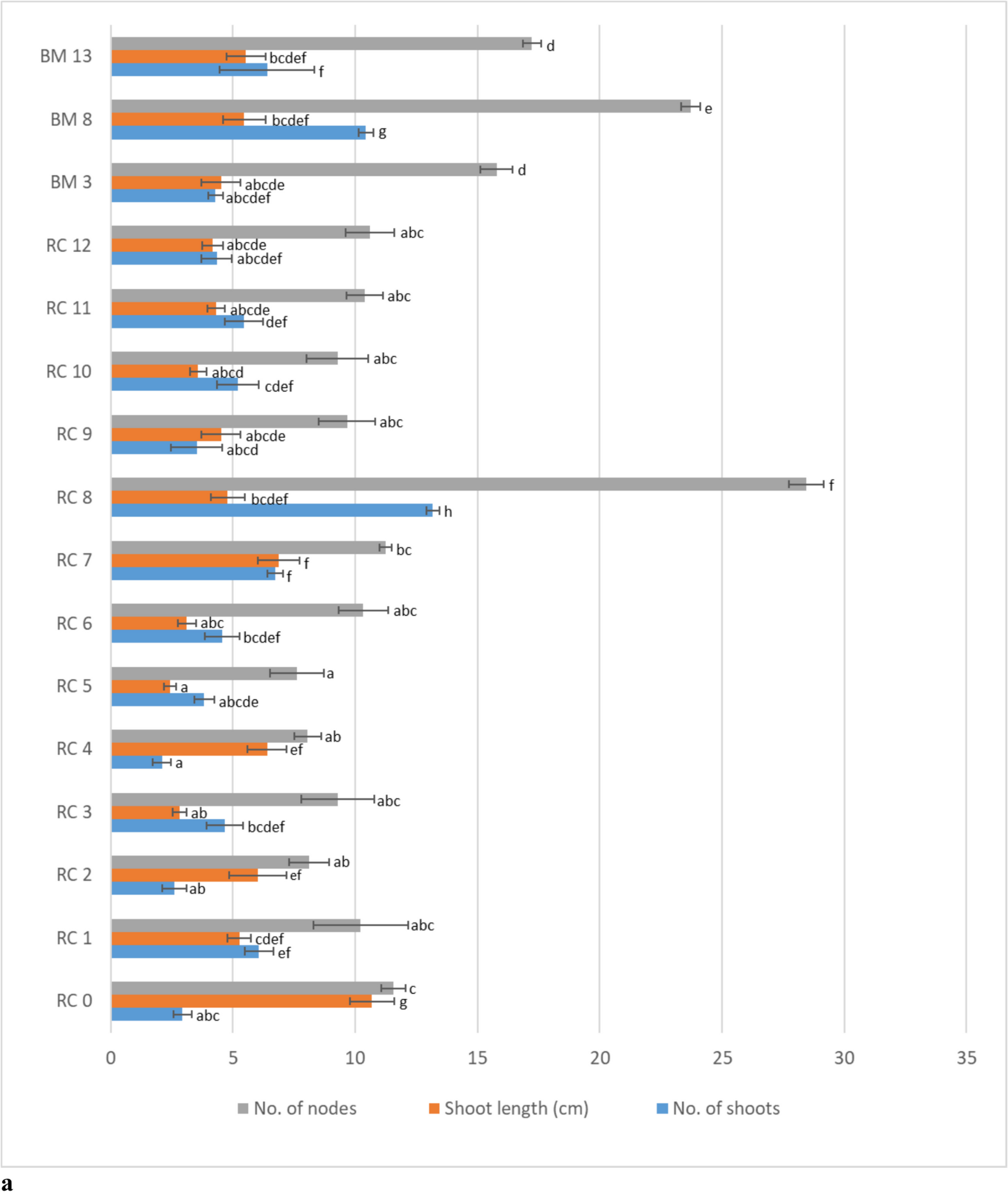
Explant cultureDifferent concentrations of cytokine and auxin (BAP and NAA; were used in the MS medium to determine the optimum media composition for callus and shoot induction from B. nigra L. After callus and shoot induction, in vitro cultured Brassica nigra explants were analyzed. Hypocotyl cultures grown on CSIM1 (Callus and Shoot Induction Medium 1), CSIM2, CSIM3, CSIM4, and CSIM5 media showed high-frequency callus formation (98.75, 94.53, 93.22, 91.22, and 92.33%, respectively) (Table 2, Fig. 1a) but no direct or indirect shoot regeneration was observed. Hypocotyl explants of Brassica juncea genotypes RH-406 and RH-555 showed shoot regeneration when supplemented with 0.2 mg L−1 NAA and 1.0 mg L−1 BAP (Kamboj et al. 2016). However, no shoot regeneration was obtained from hypocotyl explants in the present study. Cultural conditions, as well as genotype, could have a significant impact on shoot regeneration frequency (Kamal et al. 2020). Indeed, Bano et al. (2010) and Farooq et al. (2019) have reported that regeneration in Brassica highly depends on the genotype. It was observed that all calluses died in 7 to 9 d on MS medium containing only cytokinin (CSIM6 (2.0 mg L−1 BAP) and CSIM7 (4.0 mg L−1 BAP) or high concentration of cytokine compared to auxin (CSIM8 (6.0 mg L−1 zeatin,0.1 mg L−1 NAA, and 2.5 mg L−1 AgNO3. There was not any shoot formation observed from hypocotyls culture on other mediums (CSIM9 (1.0 mg L−1 BAP and 0.5 mg L−1 GA3, CSIM10 (3.0 mg L−1 BAP, 0.15 mg L−1 NAA, and 2.5 mg L−1 AgNO3), CSIM11 (2.0 mg L−1 BAP and 0.02 mg L−1 NAA). The calluses on these media survived for up to 2 mo without shoot formation. The individual application of cytokine or high concentrations of cytokine and low concentrations of auxin might have caused some toxic effects on callus cells. It is reported that auxins restrict the cytokinins effect when given at low concentrations. Conversely, high concentrations of cytokinins counteract this inhibitory effect and ultimately suppress auxin signaling (Talukdar et al. 2022). This challenge of optimizing the cytokinin-to-auxin ratio is common in plant tissue culture and can affect the efficiency of shoot regeneration (Sims 2022; Talukdar et al. 2022). The toxic effects observed could hinder the consistent production of healthy shoots, limiting the system's applicability.
Figure 1.
Regeneration of Brassica nigra Diyarbakır Ecotype from callus: Callus formation from hypocotyl explants in CSIM1 medium augmented with 0.2 mg L−1 NAA and 1.0 mg L−1 BAP (a) apex explants in CSIM2 media containing augmented with 0.5 mg L−1 NAA and 0.5 mg L−1 BAP (b), shoot explants in CSIM1 and (c) CSIM2 (d). Indirect shoot initiation from the apex callus in CSIM1 medium after callus formation, 7th d (e), 15th d (f), 19th d (g), 25th d (h). Direct shoot formation from the apex callus in CSIM1 medium (i). Multi shoot regeneration from apex explants on CSIM1 (j), CSIM3 (k) and CSIM4 medium (l). Indirect rooting was initiated after 7 d of inoculation on RIM1 media containing augmented with 1.0 mg L−1 NAA in vitro regenerated shoots (m), RIM2 media containing augmented with 1.0 mg L−1 IBA, 3rd d (n) In vitro rooting on hormone-free RIM5 medium, 20th d (o). Rooted plantlets after 30 d of inoculation on hormone-free RIM5 rooting media (p). After root formation, regenerated plants were transferred to pot culture in a growth room, Torph, 5th d (q), 13th d (r), and 25th d (s). Full plantlets of Brassica nigra transferred to 1:2 soil and torph in plastic pots (t) (Bar length is a—p: 1 cm, q—t: 5 cm). NAA, 1-naphthaleneacetic acid; BAP, 6-benzylaminopurine; IBA, indole-3-butyric acid.
Table 2. Callus and direct-indirect shoot regeneration frequency (%) from shoot, apex and hypocotyl explants of Brassica nigra L. (mean ±SE)Most of the shoot and apex explants subjected to different NAA and/or BAP concentrations showed direct regeneration (CSIM1-5) (Table 2, Fig. 1i). Shoot explants regeneration from nodes between 47.50 and 76.73% (n = 15, Table 2). Callus formation was observed on wounded parts of shoot explants in the direct regeneration period (Fig. 1c, d). Callus ratios were 88.49 and 95.18% in the CSIM4 and CSIM5 medium in the direct regeneration period, and 90.43 and 95.00 in the CSIM2 and CSIM3 medium, respectively, after direct regeneration (Table 2, Figs. 2 and 3).
All media used for apex (CSIM1-5) shown in Table 1 stimulated direct shoot regeneration from these explants (Fig. 1i). Direct shoot regeneration rates ranged from 74.07 to 100.00%. In comparison, indirect shoot regeneration rates ranged from 20.00 to 67.50% (Table 2, Fig. 3). Similar to the shoot explants, callus formation was observed on the wounded parts of apex explants. Following direct regeneration, the success rate ranged from 73.75 and 95.00% (Table 2, Fig. 1e, f, g, h). After 8 to10 days, callus formation was observed, and indirect shoot regeneration occurred with high efficiency on CSIM1 (67.50%) (Fig. 1j), and lower efficiency on CSIM3 and CSIM4 (40.00 and 20.00%, respectively) (Table 2, Fig. 1k, l). The shoot and apex explants observed direct regeneration at 7 to 10 d.
Figure 2.
The beginning of flowering in the soil and torph (1:2) (a) Flowering, approximately 4 wk (b) approximately5 wk (c) seed formation from B. nigra after about 2 mo (d) (Bar length is a: 5 cm, b—c: 1 cm, d: 1 cm).
Figure 3.
Two-way Analysis of Variance (ANOVA) test result for callus formation (a), direct shoot regeneration (b), and indirect rooting frequency (c) of explants in different Murashige and Skoog media. Vertical bars represent ± S.E. Asterisks indicate that mean values are significantly different between groups (****, p-value < 0.0001; ***, p-value < 0.0002; **, p-value < 0.0021; *, p-value < 0.0332).
The 15 to 20-day-old shoots from direct and indirect regeneration were sub-cultured to five different MS media without plant growth regulators or containing different concentrations of IBA, NAA, or a combination of both, and AgNO3 for root formation. Roots were fully formed within 5 to 20 d (Fig. 1m). Regenerated plantlets grown on RIM1, RIM2, RIM3, RIM4, and RIM5 showed 68.54, 92.27, 83.23, 58.33, and 85.71% root formation, respectively (Table 3, Fig. 3). The findings indicated that indirect root regeneration occurred only in certain types of media, and the efficiency varied. Specifically, RIM5 displayed a shorter regeneration period for indirect rooting (in 5 d) (Fig. 1o, p). This variability suggests that the system may need further optimization to consistently produce roots, a critical step for successful plant establishment and subsequent phytoremediation applications. In the media RIM2, RIM4, and RIM5, indirect root regeneration formed from the shoot. However, no indirect root regeneration was obtained from plantlets grown in RIM1 and RIM3 media (Fig. 1m, n, o). The present results show that RIM2 and RIM5 are the most suitable media for indirect rooting.
Table 3. Indirect rooting formation frequency (%) from regenerated plants of Brassica nigra L. (mean ±SE)After root formation, plants were transferred to pot culture in the growth room (Fig. 1q). Whole seedlings of B. nigra were first transferred to compost in plastic pots (Fig. 1r, s). Following 20-d of acclimatization in the compost, the plants were transferred to the soil-compost mixture at a ratio of 2:1 and were allowed to grow for flowering and seed formation (Figs. 1t, 2a, b, c). The first seeds were obtained from B. nigra Diyarbakır ecotype about 80-d after transplantation (Fig. 2d).
These results agree with those reported for other species, such as Brassica juncea and Brassica oleracea (Mollika et al. 2011; Gerszberg et al. 2015). Moreover, under the experimental conditions of the current study, increasing IBA concentration promoted both rooting and root numbers but prevented root elongation. This trade-off between root initiation and elongation is a critical factor to consider, especially for plants intended for phytoremediation, where root depth and spread are important for metal uptake. Similar results have been shown with other plant species when supplied with IBA (Al-Jaboory et al. 1998; Kotsias and Roussos 2001).
Currently, a limited number of studies on regeneration of different ecotypes of Brassica nigra are reported. For example, Das et al. (2010) reported successful hairy root regeneration from B. nigra (genotype B-54), which is the most common oilseed crop in India. Hairy root cultures are advantageous for their high growth rates and genetic stability, making them suitable for phytoremediation studies. Compared to the current in vitro regeneration system, hairy root cultures may offer a more efficient method for producing biomass and studying metal accumulation. In addition, Gu et al. (2014) developed efficient microspore embryogenesis from B. nigra (genotype B-431), which provides a haploid plant production method. This approach holds significant potential for genetic studies and breeding programs to improve phytoremediation traits. However, the in vitro regeneration system described in this study lacks essential genetic tools, potentially limiting its efficacy in comprehensive genetic improvement endeavors. The B. nigra Diyarbakır ecotype used in this study is a Cu-tolerant and accumulating plant native to the Southeastern Anatolian region. Therefore, it is considered a unique Cu accumulator ecotype with potential for further evaluation in the phytoremediation of Cu-contaminated soils. To the best of current knowledge, this is the first study on the B. nigra Diyarbakır ecotype that compares the effects of different plant growth regulators, such as BAP, NAA, and IBA, on callus, shoot, and root regeneration from three types of explants; apex, shoot, and hypocotyl. The results show that it is possible to obtain formation from hypocotyl and direct or indirect shoot formation from apex in B. nigra Diyarbakır ecotype. On the other hand, direct orindirect shoot formation was obtained only from the apex and stem in this ecotype.
Metal tolerance assayUnderstanding the details of the uptake, transportation, and detoxification/exclusion mechanism of essential or toxic mineral elements and organic contaminants through biochemical, physiological, and molecular approaches is a prerequisite for improving phytoremediation technologies. In other words, the more detailed description of events taking place during the uptake of mineral elements to roots, transportation to stem, leaves or seeds, storage or exclusion will pave the way for the development of better methods for phytoremediation (Dalvi and Bhalerao 2013; Komal et al. 2015; Pasricha et al. 2021). For this purpose, the Cu tolerance level of the B. nigra Diyarbakır ecotype was screened. The Cu accumulation capacities of different organs of B. nigra Diyarbakır Ecotype plants regenerated from tissue culture were determined both in MS medium and soil culture.
Cu accumulation capacity of B. nigra Diyarbakır ecotype grown in MS mediumB. nigra Diyarbakır Ecotype regenerated from tissue culture (30-d after the shoot and root generation) was taken into MS media containing different concentrations of Cu (Fig. 4). The plants were exposed to 50.0 to 1000.0 μM of Cu2+ for 72 h. It was observed that the Diyarbakır ecotype survives in MS medium containing up to 1000.0 µM Cu. It has been known that plants exhibit sensitivity to elevated levels of Cu and display toxic symptoms when exposed to excessive amounts (Saleem et al. 2020; Mir et al. 2021a). Plants experience morphological alterations, disruptions in metabolism, modifications in biochemical processes, and inhibited growth due to these symptoms at higher concentrations. It has been reported that Cu has a toxic effect on some species, even at low concentrations. Quartacci et al. (2003) investigated the uptake and transport of Cu in four different Brassica species. They observed Cu toxicity effects in some species at low concentrations (5 µM). The current study showed no morphological difference between controls and plants treated with 50.0 to 1000.0 μM Cu (Fig. 4a).
Figure 4.
Thirty-d-old B. nigra plants were transferred to Murashige and Skoog media containing different levels of CuSO4, Control (a), 50.0 µM (b), 100.0 µM (c), 200.0 µM (d), 500.0 µM (e), 1000.0 µM (f) (A) Metal uptake capacity of whole Brassica nigra Diyarbakir ecotype plant (µg g−1 D.W.) * D.W. = Dry Weight. Vertical bars represent ± S.E. Asterisks indicate that mean values are significantly different between groups (****, p-value < 0.0001; ***, p-value < 0.0002; **, p-value < 0.0021; *, p-value < 0.0332) (B) (Bar length is 5 cm).
The plant's roots and shoots (stems and leaves) were analyzed separately for Cu accumulation. The results were calculated according to the dry weights of the plants and were given in µg g−1. The highest total Cu accumulation capacity was found in plants treated with 1000.0 μM CuSO4, reaching 8347.0 µg g−1 (Fig. 4b). In contrast, only trace amounts of Cu, around 98.0 µg g−1, were detected in the control plants. The amount of Cu in the tissues increased in parallel with the rising Cu concentration in the medium. It was observed that the main accumulation of Cu in the plant was in the shoots, which indicates the efficient translocation of Cu from roots to shoots in this ecotype. A previous study found that different B. nigra ecotypes accumulated Cu in their root tissues when exposed to 25 µM Cu in hydroponic culture. It was also determined that high Cu concentrations (50.0 to 500.0 µM) increased the Cu content in both root and shoot of this ecotype after a 72-h treatment (Cevher-Keskin et al. 2019). This research also showed that Cu accumulated mainly in the shoots of the plants grown in the MS medium (Fig. 4b).
Cu accumulation capacity of B. nigra Diyarbakır ecotype grown in soil cultureCopper is a vital element that plays a significant role in many essential physiological activities of plants. It acts as a catalyst in redox reactions, contributing to numerous metabolic processes necessary for plant growth and development (Mir et al. 2021a, b; Chen et al. 2022). However, excessive accumulation of Cu in plant tissues can lead to toxic effects and become lethal (Yruela 2009). Cu exists in various forms in the soil, but it is readily taken up and metabolized by plants as Cu2+ (Rusjan 2012). The Cu content in the soil generally ranges from 2.0 to 250.0 μg g−1, and Cu-tolerant plants can accumulate around 20.0 to 30.0 μg g−1 DW (dry weight) of Cu (Sam and Eruni 2022). The uptake, absorption, and accumulation of Cu from the soil primarily depend on its concentration in the soil, the duration of exposure, and the different genetic structures of the species. For example, Mir et al. (2021b) reported maximum accumulation in the roots compared to the shoots in B. juncea grown at different Cu concentrations in the soil. Mahmud et al. (2013) showed that plants belonging to the Poaceae family mainly accumulated Cu in both the roots and shoots. Buddleja nivea accumulated more Cu in its roots, followed by the leaves and stems (Rehman et al. 2019a).
In the current study, the plants propagated in tissue culture were transferred to the soil, as described in the materials and methods Section 30-d-old plantlets were used for Cu treatment. As the plants have shown very high tolerance to the Cu in MS medium, 200.0 and 500.0 ppm CuSO4 were selected for soil application. The plants were exposed to Cu for 3 wk until fully grown to be able to analyze the different tissues of the plant. The translocation rate of Cu from root to shoot is quite slow as Cu is retained in the root apoplast. Recently, Ambrosini et al. (2018) conducted a study on the translocation of Cu in 'Red Niagara' (Vitis labrusca), a well-known grape variety in Brazil. The findings suggested that grapevines respond to Cu stress by accumulating Cu in the apoplast and limiting its translocation to the shoots.) Zhou et al. (2007) also demonstrated that Cu adsorption in maize root cell walls was so tight that many metal ions were retained in the root apoplast. It was observed that as growth progresses, there is an increase in Cu accumulation, with maximum accumulation in roots and leaves compared to shoots. The Cu concentration varied between 69.0 to 1525.0 µg g−1 in the roots and 51.0 to 939.0 µg g−1 in the leaves. Similarly, Rather et al. (2022) reported the highest Cu accumulation in both roots and leaves in B. juncea cultivar Pusa Tarak, regardless of Cu concentration. In support of these results, no toxic symptoms were observed on Diyarbakir ecotype plants in either Cu concentration at the end of the 3 weeks in soil culture. All plants were flowering at the end of 3 wk (Fig. 5a). The accumulation of Cu in flowers and shoots was minimal compared to that in roots and leaves, which supports the findings of Gonzaga et al. (2018), who also investigated the Cu uptake capacity of B. juncea in Cu-contaminated soils. It is noteworthy that at 200.0 ppm Cu, leaves accumulated the highest amount of Cu (601.0 µg g−1), whereas, at 500.0 ppm Cu, roots accumulated a higher concentration of Cu (1525.0 µg g−1) (Fig. 5b). These results indicate an upper limit for Cu transport to the leaves, and then Cu starts to accumulate in the roots.
Figure 5.
Different CuSO4 concentrations were treated to Brassica nigra L. Diyarbakir ecotype in soil culture (Control, 200.0 ppm, 500.0 ppm µM CuSO4) (a) Metal uptake capacity of B. nigra L. Diyarbakir ecotype in soil culture (µg g−1 D.W.) * D.W. = Dry Weight. Vertical bars represent ± S.E. Asterisks indicate that mean values are significantly different between groups (****, p-value < 0.0001; ***, p-value < 0.0002; **, p-value < 0.0021; *, p-value < 0.0332) (b) (Bar length is 20 cm).
Similarly, it has been reported that the translocation of Cu towards the aboveground plant parts in Fallopia convolvulus is effectively restricted by the significant Cu accumulation in the roots (Pedersen et al. 2000). Similar results were also obtained in Oryza sativa, Triticum aestivum, and Boehmeria nivea L (Thounaojam et al. 2012; Mahmud et al. 2013; Rehman et al. 2019b). These studies demonstrate that Cu tends to accumulate in the root tissues and has limited movement toward the aboveground organs (Cevher-Keskin et al. 2019).
Comments (0)