
Remember me


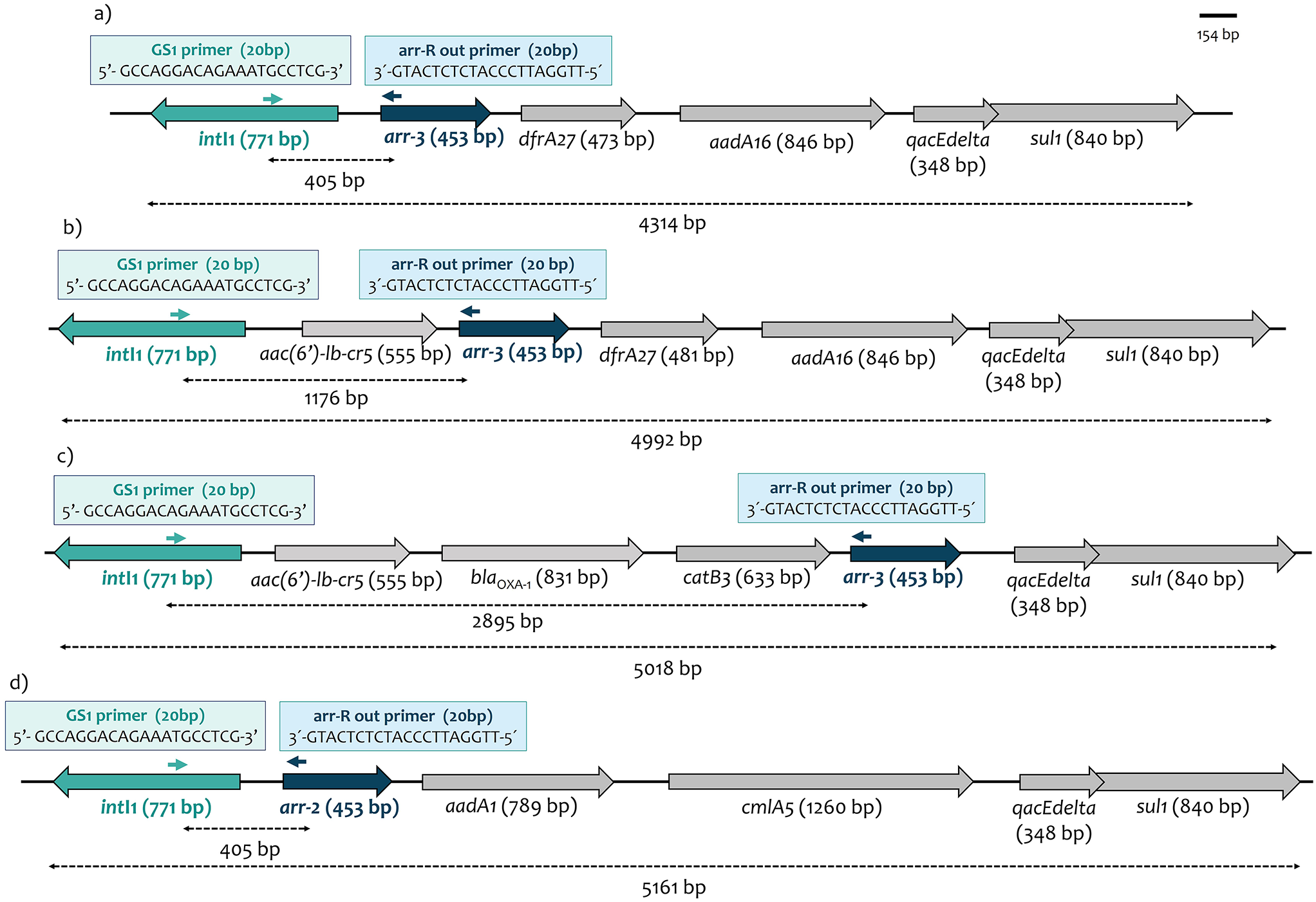

Seven individuals were chosen for the RD group regarding the absence of antibodies against all serotypes of DENV (Fig. 1A). The group of SD was constituted of seven anti-DENV seropositive individuals. The sex ratio was the same in both groups and average ages of 28 years in SD and 23 years in RD group (p = 0.057, range = 21–34 years), (Fig. 1A).
Fig. 1
Phenotypic/immunological data of individuals, and viral titration of their monocytes after DENV-2 infection. (A) Gender, age and DENV antibodies detection by hemagglutination inhibition in the 2 groups. (B) Log10 viral titer median on days 1, 2, 3, 4, 5 post infection. Error bar figures inter-quartile range. Statistics (Mann-Whitney test) shows significant difference on days 1, 2 and 3 with p-value = 0.0006 (***) but no significant difference on days 4, 5
To investigate differential susceptibility to infection between the 2 groups, donor monocytes were inoculated with DENV-2 at MOI 1 pfu/cell. The efficiency of replication was monitored within 5 days post-infection, showing a mean viral titer in the SD group 9-fold higher than the RD group at 1-day post-infection (dpi), 13-fold higher at 2 dpi, and 5-fold higher at 3 dpi (Fig. 1B). Therefore, monocytes from RD participants yield a reduced viral progeny.
Some miRNAs show differential expression in the context of DENV resistanceTo explore the miRNA role in the resistance to DENV we performed a miRNA chip hybridization assay for mock-infected cells and for DENV-infected cells in both RD and SD groups. After data quality control two samples were removed, one from RD mock-infected cells due to poor hybridization and one outlier from SD infected group. We performed a pairwise comparison of miRNA expression between the experimental groups and miRNA were considered differentially expressed (DE) with absolute values of fold change (FC) > 1.25 and q-value < 0.05. The list of DE miRNAs in each comparison are given in supplementary Tables ST1, ST2, ST3, and ST4.
First, comparing DENV-infected vs. mock-infected monocytes in the SD group we found seven miRNAs, including miR-103b-1, miR-6879-3p, and miR-550b-1, that may participate in the cell response to infection process (Fig. 2A, Supplementary Table ST1), ruling them out as exclusive mechanisms of resistance in the RD group. Secondly, we evaluated the alteration of miRNAs expression upon infection in monocytes from RD group. We found 24 downregulated and 10 upregulated miRNAs in DENV-infected cells, including miR-576-3p (FC=-2.6, q-value = 8.0 × 10− 3) and a few members of miR-548 family (548n, 548b-5p, 548j-5p, 548i) among the most repressed ones. Conversely, miR-501 (FC = 1.73, q-value = 2.7 × 10− 2), miR-576-5p (FC = 1.35, q-value = 1.5 × 10− 2), and miR-155 (FC = 1.31, q-value = 9.37 × 10− 3) were upregulated in DENV-infected monocytes of RD participants (Fig. 2B, Supplementary Table ST2). This miRNA expression pattern observed in RD could play a role in the resistance mechanisms.
Fig. 2
Representation of differentially expressed miRNA. (A to C): Volcano plots showing changes in miRNA expression levels according groups (RD = Resistant to dengue viral infection, SD = Susceptible to dengue viral infection). Red dots denote significantly differentially expressed miRNA (q-value < 0.05, Log10 q-value > 1.3) whose expression changed by more than 1.25-fold. Vertical line represents log2FC ± 0.32 threshold, and horizontal dotted line represents statistical significance threshold (q-value = 0.05). (D) Overall view of expression of selected miRNA. Salmon boxes represent overexpression while blue boxes mean reduction of expression. In the center of each colored box is the Fold Change (+/-) N.S. Not significant: q-value ;> 0.05
Subsequently, we studied the differences between infected monocytes from RD group vs. SD group, as this comparison shows DE miRNAs that may participate in viral blocking pathways present in RD individuals (Fig. 2C, Supplementary Table ST3). We found 19 downregulated and 23 upregulated miRNAs in RD, with a remarkable 5-fold increase in the expression of miR-132-3p (FC = 5.0, q-value = 2.8 × 10− 2). Among upregulated miRNAs, it is worth noting two miRNAs described in the literature for their role in immune response, let-7e-3p (FC = 1.33 q-value = 4.7 × 10− 2) and miR-155 (FC = 1.29, q-value = 1.0 × 10− 2). In addition, miR-576-5p (FC = 1.27, q-value = 3.6 × 10− 2) and the aforementioned miR-155 are upregulated as in the DENV-infected vs. mock-infected cells comparison in the RD group (Fig. 2B), suggesting a protective role in DENV infection. Additionally, we obtained 17 DE miRNAs comparing mock-infected monocytes from RD to SD groups (Supplementary Table ST4), to find some miRNAs that could play a role in the resistance to the infection in the monocytes basal state. We found that miR-576-3p (FC = 2.23, q-value = 1.7 × 10− 4) along with three members of miR-548 family (548i, 548au-5p, and 548b-5p) are overexpressed in RD.
Finally, we summarized the most interesting DE miRNA based on the FC in three comparisons, including published studies that support a potential role in the pathogenesis of infectious diseases and antiviral immune response (Fig. 2D). Therefore, we chose miR-132-3p, miR-576-3p, and miR-155-5p as candidates for performing functional assays and assessing their role as antiviral miRNAs. In the case of miR-155, we selected the − 5p arm since it is preferred in the biogenesis process and regulate up to 220 gene targets defined in miRTarbase (4 gene targets for − 3p arm).
Mir-155-5p upregulation decreases DENV infection in HeLa cellsWe performed functional assays with the selected miRNA candidates to investigate their effect in the infection and viral production. For this purpose, we performed a transient upregulation of miR-132-3p, miR-576-3p, and miR-155-5p through the transfection of mimics of each miRNA. The transient upregulation of miR-155-5p caused a significant reduction in infection rate, although miR-576-3p and miR-132-3p overexpression did not affect intracellular detection of DENV E protein (Fig. 3A). These results are congruent with the more than one log10 reduction in viral titer upon miR-155-5p mimic treatment (Fig. 3B).
To verify if miR-155-5p upregulation produced an antiviral effect with all DENV serotypes, we infected with DENV-1, DENV-2, DENV-3, and DENV-4 mimic-transfected HeLa cells, either with miR-155-5p mimic or a scramble miRNA (mimic negative control). We observed a reduction in the rate of infection 48 h post-infection (hpi) of 36% in DENV-1, 62% in DENV-2, 26% in DENV-3, and 68% in DENV-4 upon overexpression of miR-155-5p (Fig. 3C). The reduction of viral titers supports the data of infection rates, decreasing a 92% for DENV-1, 79% for DENV-2, 75% for DENV-3 and 85% for DENV-4 48hpi, suggesting that the overexpression of miR-155-5p elicits an antiviral response in HeLa cells.
Fig. 3
Functional study to show effect of miRNA mimic transfection on DENV infection. (A) Cytometry on DENV-1 infected HeLa cells, 48 h post infection, transfection 24 h, and (B) viral titration of supernatants of those cells. (C) Cytometry on DENV-1, DENV-2, DENV-3, DENV-4 infected HeLa cells, 48 h post infection, transfection 24 h. (D) Viral titration of supernatants of those cells. Statistics: Mann-Whitney non-parametric tests ** p = 0,0079; * p = 0,0159
Mir-155-5p shapes transcriptomic landscape restricting dengue pathogenesisWith a view to describe what processes and genes could be altered by miR-155-5p upregulation, we performed an RNA-Seq on Hela cells 24 h after mimic transfection. We identified in miR-155-5p mimic transfected cells 265 downregulated DEGs (FC<-2, FDR < 0.05) and 112 overexpressed DEGs (FC > 2, FDR < 0.05) (Supplementary Table 5), of which 260 DEGs were defined as miR-155-5p targets in miRNA-mRNA interaction databases (Fig. 4A). Of those, 209 genes are downregulated and presumably under the direct repression brought on by mir-155-5p overexpression. The over-representation analysis showed that a total of 115 (FDR < 0.05) biological terms from the Reactome database are enriched with the 260 DEGs targeted by miR-155-5p (Fig. 4B, Supplementary Table 6), and thus influenced by miR-155-5p upregulation. These genes controlled by miR-155-5p are involved in immune response processes, transcriptional activities, the regulation of TP53 signaling, in vesicle and molecule trafficking and cell cycle regulating-signaling (MAPK, EGFR, BRAF, AKT) pathways. In addition, those DEG participates in processes regulated by interferon signaling, endocytosis, apoptosis, or mTOR pathway (Supplementary Table 6). It is compelling that 18 DEGs targeted by miR-155-5p are described as interactors of DENV proteins (Fig. 4C). Downregulated genes (ANXA2, PEPB1, TNFRSF10B, RPL17, HSD17B12) are explained because of direct repression of miR-155-5p, although others such as HMOX1, MT2A, TRAFD1 or EIF4G2 could be overexpressed by an indirect mechanism of upregulation by this miRNA.
Fig. 4
RNA-Seq in HeLa cells transfected with miR-155-5p mimic or scrambled mimic. (A) Venn diagram representing DEG in Hela cell transfection (pink), DENV-Human protein interactions registered in DENVHunt Database (grey) and experimentally validated targets of miR-155-5p from miRTarbase and Tarbase (yellow). (B) Bubble plot representing biological terms from Reactome enriched with the 260 DEGs targeted by miR-155-5p as registered in miRTarbase and Tarbase. The 30 terms with lower FDR are shown. Bubble size refers to the number of genes that compose the biological term. Chromatic scale indicates the percentage of genes that are DEG in each biological term. Transcript. = Transcription. (C) Heatmap of the 18 DEG in Hela cell transfection experiment defined as experimentally validated targets of miR-155-5p and that interact with DENV proteins. Chromatic scale represents Z-score of normalized counts
Comments (0)