
Remember me



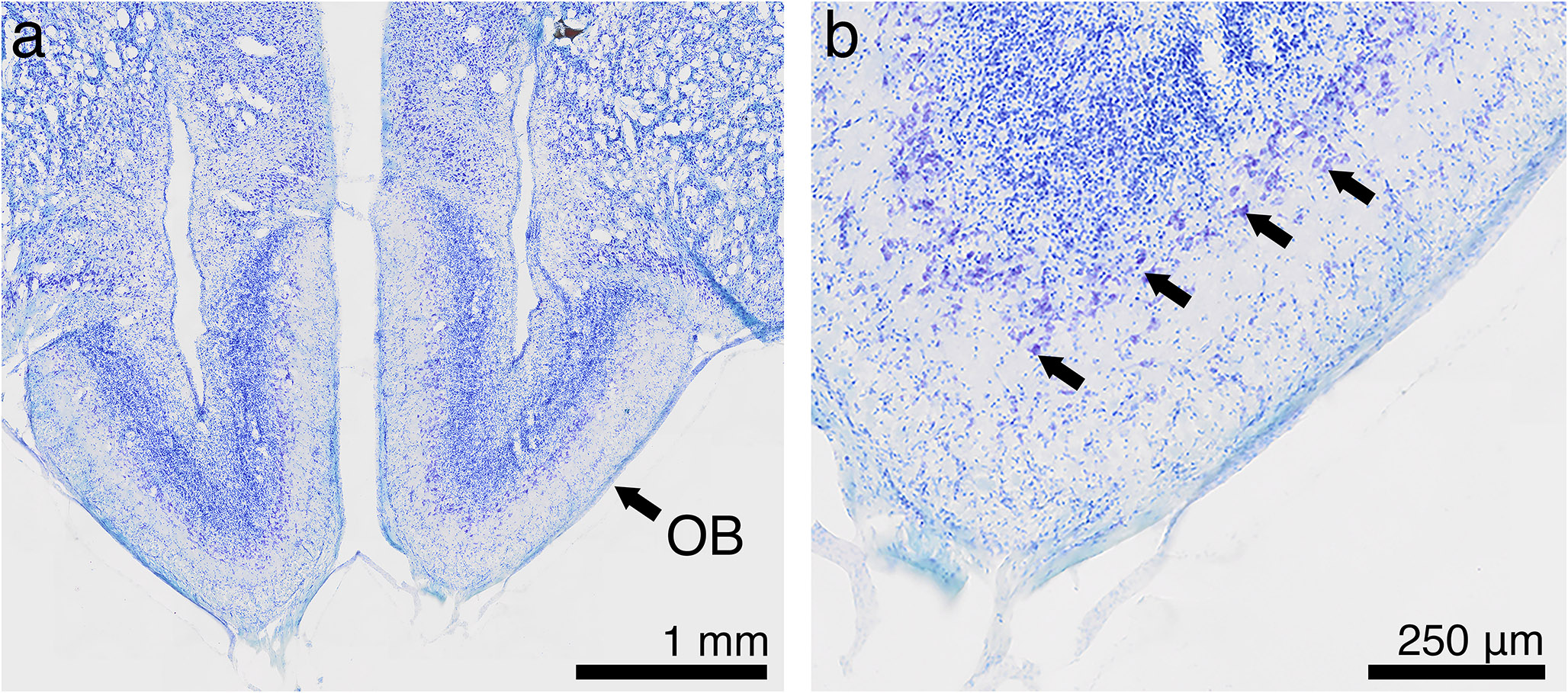

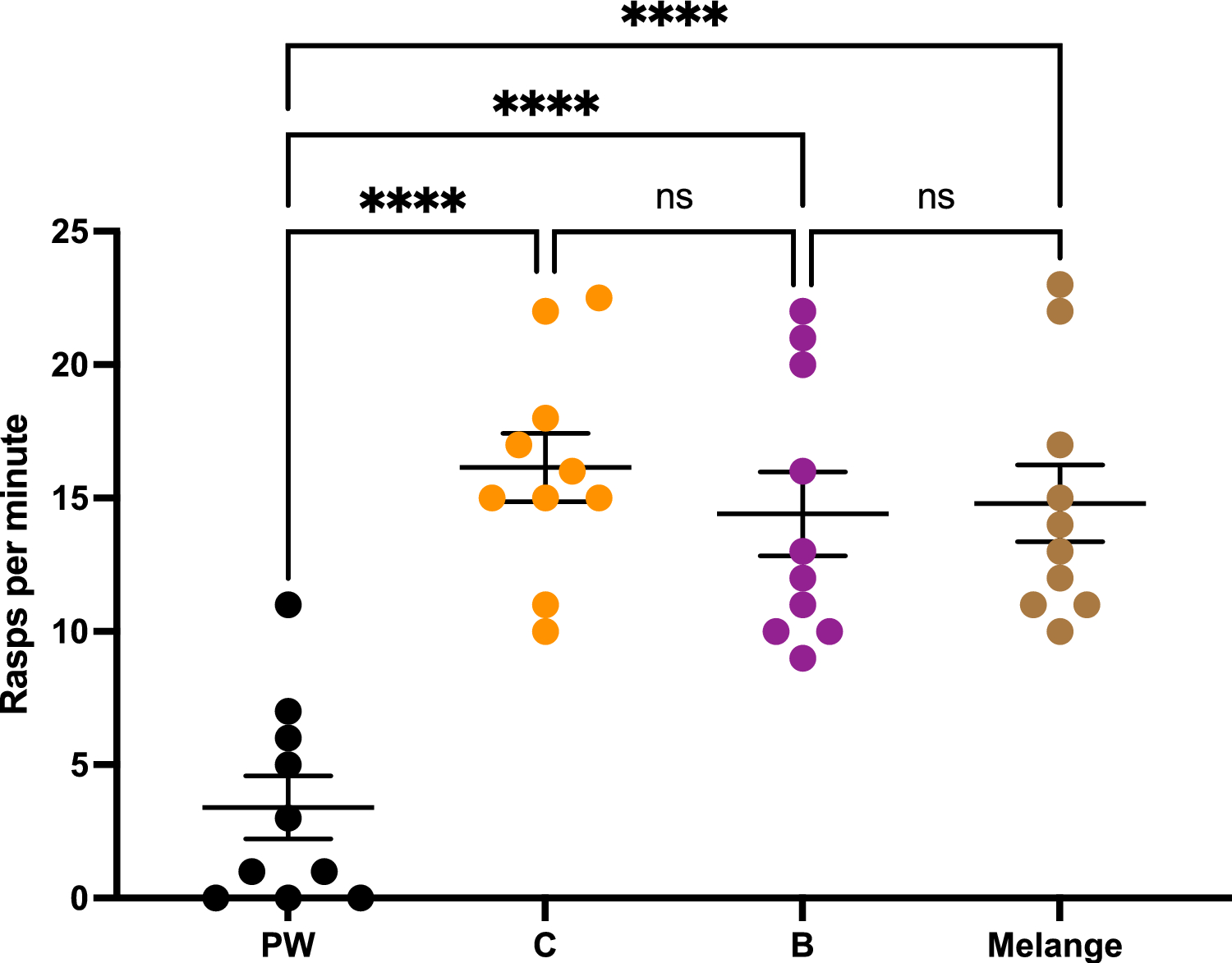

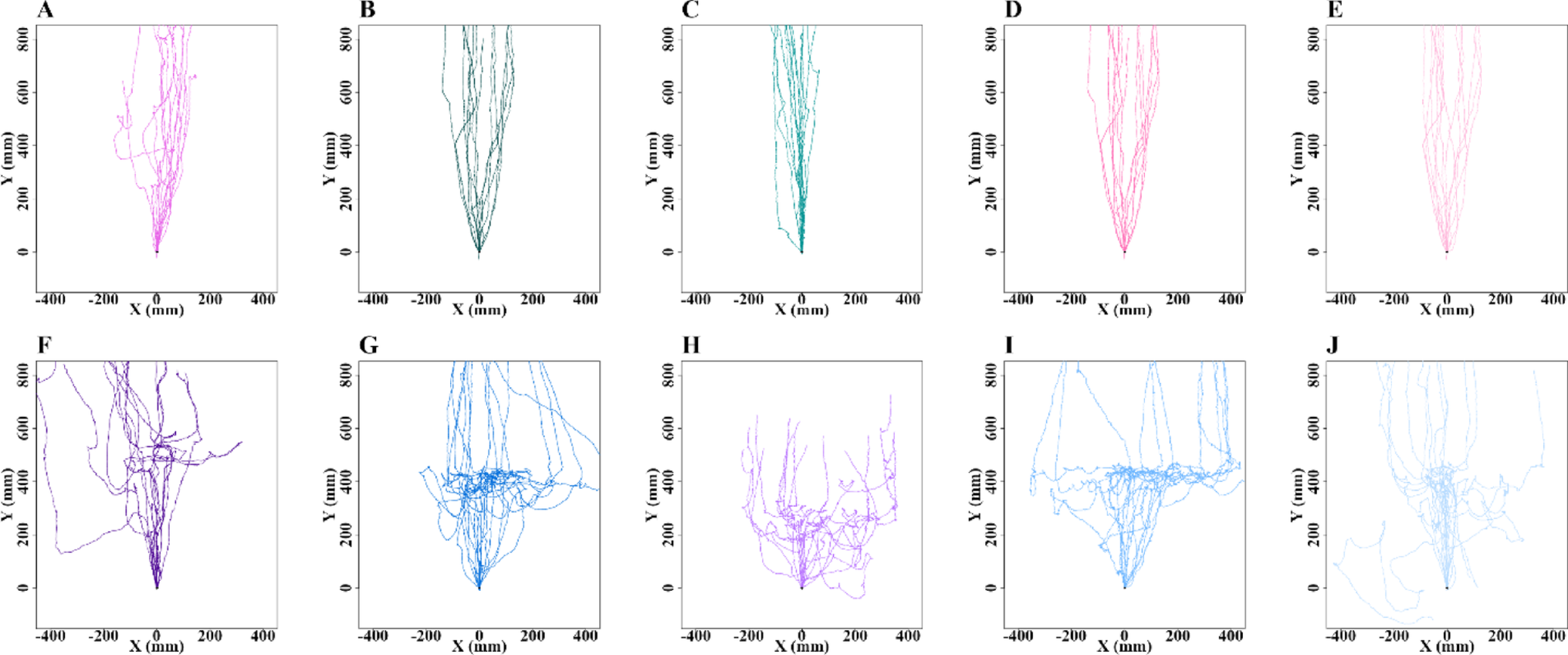
Cooperative foraging behavior is thought to require social exodigestion of food, and therefore likely requires functional salivary glands. To test the role of salivary glands, clustering assays were performed (Dombrovski et al. 2017). One wild-type strain (CS) and three transgenic lab strains (P, sgs, tai, see methods) were used. In addition, a blind GMRhid and the cross of sgs and tai, sgstai (see methods) were also tested. Blind GMRhid larvae are very inefficient at clustering due the requirement of vision for this synchronized behavior (Dombrovski et al. 2019). The sgstai larvae are expected to have non-functional salivary glands due to the overexpression of the metamorphosis-inducing gene taiman, tai (Farkaš et al. 2014). In comparison to wild type, sgstai larvae have reduced salivary glands (Fig. 1a). Initially, the apoptosis-inducing gene hid (see methods) was overexpressed in salivary glands but this proved mostly lethal at the pupal stages. Salivary glands produce a glue which allows pupae to stick to the vial sides, an event which did not generally occur in sgsGal4/UAS-hid. Therefore sgsGal4/UAS-tai (sgstai) was used as a salivary gland depleted animal. Each of the 6 strains were first tested for clustering in a 2D assay (Dombrovski et al. 2017). Pre-processed food was placed between two glass slides, and the 40 clustering larvae from a crowded vial were loaded and video documented for 24 hours. Clustering was measured as the proportion of larvae in clusters as described (Dombrovski et al. 2017). Clusters were defined at 4 or more synchronized larvae in the food. The clustering proportion was averaged from 240’, 360, and 480’. About 50% of larvae were in clusters for the wild type and 3 lab transgenic strains (Fig. 1b). Few of salivary gland depleted sgstai clustered while loss of vision (GMRhid) halved clustering rates as described before (Dombrovski et al. 2019). The sgs and tai are the parental strains for sgstai and this shows that salivary glands are likely a key part of clustering.
Fig. 1
Allee analysis of fitness in lab strains. (a). Salivary glands of sgs (sgsGAL4) and sgstai (sgsGAL4/UAStai) in the first 24 h of the 3rd instar. Salivary gland cells are visible in sgs but not in the sgstai samples. Sale bar is 200 μm. (b). 40 larvae from the 6 strains used in this study were placed in a 2D clustering assay. The average number of larvae in clusters from each assay was averaged over several experiments and indicated. About half of the larvae are in clusters for the 4 normal strains but greatly reduced in blind larvae and almost absent in cheater sgstai. Values are expressed as the average of multiple assays and the standard error of the mean. The number of samples is shown for each data point. Significance, after normality test, is based on comparison to CS and ANOVA analysis followed by Tukey’s Test. * p < 0.05; **** p < 0.0001. Probabilities for non-significant differences are shown. (c-i). Various numbers of L2 larvae were added to pre-processed vials and raised to adults. The size of the female wing was used as a fitness measure. For CS, tai, sgs, P strains there is a peak or plateau point in wing size at about 40 larvae per vial. Placing CS in the dark, where clustering is inefficient results in a steady decay curve likely due to competition. Indicated are the averages and standard deviation as errors. The number of samples is shown for each data point. Significance, after normality testing, is based on comparison to wing size at 10 animals and ANOVA analysis followed by Tukey’s Test. * p < 0.05; ** p < 0.01; *** p < 0.001. **** p < 0.0001. Probabilities for non-significant differences are shown
Cooperative foraging behavior should also exhibit an Allee effect in terms of the number of larvae in a defined substrate. There is expected to be an optimal number of cooperating larvae in a vial in which the positive effects of the group offsets competition. Previous studies used 200 larvae per vial (Dombrovski et al. 2017, 2020) which also likely featured significant intra-specific crowding effects (Miller and Thomas 1958; Venkitachalam et al. 2022). In order to find the optimum, a range of different numbers of second instar larvae were added to preprocessed vials as described (Dombrovski et al. 2017). Preprocessed vials have previously hosted about 100 wild type CS larvae from egg to pupal stages and about half of the about 5 ml of food was now in a semi-liquid, partially digested state (Dombrovski et al. 2017). The rationale for using these vials is that food is optimized for clustering based on physical properties, digested state and microbiome. Therefore, larvae entering these vials should be able to cluster. Larvae were loaded at the second instar stage so that they could enter the critical clustering period at the start of the third instar (Dombrovski et al. 2019). Vials were loaded with 10, 20, 30, 40, 50, or 200 larvae. The expectation is that any measure of fitness should decay with an increasing numbers of larvae due to competition but there should also be a peak in the distribution due to cooperation (Allee 1927). Larvae were allowed to develop into adults and female wing size was assessed as a fitness marker. Female wing size is directly related to body size and scales with fecundity (Gilchrist and Partridge 1999, 2001). Wing size is also directly related to how much larval clustering occurs (Dombrovski et al. 2020; Williamson et al. 2021). CS and P (Fig. 1c, d statistics Fig. 2a, f) show a fitness peak of about 40 larvae per vial. The fitness gain for 40 P larvae, over 10, remains consistent for mixes with CS, sgs, tai, sgstai but not for mixes with blind GMRhid or with any combinations in the dark (Fig. 2f). The lack of difference for fitness between 10x and 40x larvae for sgs and tai might be due to greater intraspecific competition. But for measuring optimal clustering conditions for CS and likely other strains, about 40 larvae per vial is optimal.
Fig. 2
Fitness of larval mixtures. (a-f) Wing size fitness of 40x larvae/vial grown to adult of select mixes for 6 labs strains including blind GMRhid and salivary-gland-depleted sgstai. Significance, after normality testing, is based on comparison to wing size at 10 animals and ANOVA analysis followed by Tukey’s Test. * p < 0.05; ** p < 0.01; *** p < 0.001. **** p < 0.0001. Probabilities for non-significant differences are shown. Values are expressed as the average of the indicated number of samples and the standard deviation. (a). CS: except for development in the dark, all 40x larval combinations with various amounts of the P strain show gains over 10x. (b-c). tai/sgs: These strains have no distinct Allee peak but fail to show decreased fitness as numbers grow until about 40. This gain is lost in cluster free dark-developed vials. P can substitute for either sgs or tai. (d). GMRhid (blind): No gain or loss is seen for 10-40x larvae of any mix. (e). sgstai (larval gland depleted): Gain is seen for sgstai when mixed with P either 10:30 or 30:10. This is lost when P is replaced with blind GMRhid or incubation in dark. (f). P (host strain): P strain shows a gain from 10 to 40 larvae and P can be substituted for CS, sgs, tai, sgstai but not blind GMRhid. Both P and sgstai (e) gain from mixing. (g). Survival of various sgstai mixes with P, P-dark or GMRhid compared at increasing larval density. In each case, 10 sgstai were placed in a vial in increasing numbers of hosts as indicated on the X-axis. At 30 and 40 P, there is greater survival of sgstai that with GMRhid as hosts or P in the dark. Significance, after normality testing, is based on comparisons between dark reared and use of blind GMRhid as host and calculated with ANOVA analysis followed by Tukey’s Test. ** p < 0.01; *** p < 0.001. **** p < 0.0001. Values are expressed as the average of the indicated number of samples and the standard deviation
If 40 larvae is the optimal number in a vial for the fitness benefits of clustering, can these larvae be of mixed composition? If so, it is predicted that sgstai larvae should be able to gain from a wild type host. The P strain, which has distinguishable white eyes, was used as a host for all other strains. A total of 40 L2 larvae were loaded into each vial, grown to adulthood and ha their wings measured. Either 10x test and 30x P or 30x test and 10x P were used. CS and P are interchangeable with each other (Fig. 2). 30xP substitutes for 30xCS (Fig. 2a) and vice versa (Fig. 2b). This is also true for the sgs and tai strains (Fig. 2c-d). However, this effect is lost when the vials are kept in darkness which attenuates cooperation (Fig. 2a-d). Therefore, the Allee effect gain in going from 10 to 40 larvae does not matter on composition. However, this is not true for blind GMRhid larvae. Substituting 10xP with 10xGMRhid along with another 30xP removes the Allee effect gain. Blind larvae presumably block clustering in the host by interfering with group synchronization (Dombrovski et al. 2020). sgstai larvae, which do not cluster well, and do not show an Allee effect by themselves, now show a gain in wing size when mixed with P. This is presumably due to the salivary gland-depleted strain using the salivary products of the host. However, this occurs with both 10:30 and 30:10 proportions. The expectation had been that going from 10:30 to 30:10 cheaters to host, that the gain would not be seen. In fact, the depleting effect of excess sgstai ‘cheating’ was expected to be seen in the P hosts, which should have lost their Allee effect. Instead, they show gain (Fig. 2f). Therefore, salivary gland depleted larvae show gain with a wildtype host which also gains from the mix. This gain can also be seen in adult survival (Fig. 2g). Survival decreases with increasing numbers of larvae in the vial but there is an increase in wing size at 10sgstai and 30P compared to the same blend in the dark or using GMRhid as host (Fig. 2g). Therefore, the increase in wing size matches survival.
To investigate the mutual gain between P and sgstai further, 2D clustering assays were performed with blends. 10CS&30P and P cluster at about the same amount while more larvae cluster in 10sgstai&30P. GMRhid block all clustering when mixed with 30P (Fig. 3a). Therefore, sgstai increases clustering when mixed with a wild type host. To examine inclusion, food coloring-labeled larvae were add to either CS or P clusters and the length of time spent in the cluster measured (Dombrovski et al. 2017). Transplanted CS larvae spend about an hour in either CS or P clusters as measured before (Fig. 3b). sgstai larvae spend over double this time in P clusters (Fig. 3b). To examine if this increased cluster inclusion time is based on better synchronization (Dombrovski et al. 2017), high-resolution videos were made of transplanted CS or sgstai larvae. The sgstai larvae do not synchronize with host (Fig. 3c). In previous studies, such non-synchronizing larvae did not stay long in clusters (Dombrovski et al. 2017, 2019; Williamson et al. 2021). Blind GMRhid larvae delay inter-larval movements at 0.72s+/0.06 (Dombrovski et al. 2017)which is similar to that between sgstai and P (0.72s+/-0.06) and much longer than between CS and P (0.43+/-0.06 s) reported here. To examine the clustering dynamics, 2D assays were established and every cluster within the first 6 h documented. The number of larvae in each cluster was counted and followed for its duration. The cluster size was the average number of larvae for the lifespan of the cluster. This was about 7 for all conditions except sgstai and 10GMRhid&30P. Only 3 clusters were seen for sgstai and those for 10GMRhid&30P were close to the limit for the definition of cluster, which is 4 larvae. The average size of sgstai mixed clusters was about the same as that of P and so this does not explain the increased clustering. The average cluster life span was also measured. This was larger than 10CS30P. Therefore, sgstai larvae increase overall clustering by increasing the lifespan but not the size of a cluster (Fig. 3e).
Fig. 3
Clustering dynamics of mixes. (a). The proportion of larvae clustering in 2D was measured for three key mixes: P alone, 10CS&30P, 10GMRhid&30P, and 10sgstai&30P. About half of larvae are in clusters for P, 10CS&30P. This is about the same for CS alone (Fig. 1a). GMRhid blocks all clustering in P. However, sgstai increases the amount of clustering when substituted for P or CS. All graphs in this figure show the average with standard error. The number of samples is shown for each data point. Significance was calculated, after a normality test, by ANOVA analysis followed by Tukey’s Test and compares each sample to the first one on the left side of the graph. Other comparisons are as indicated. * p < 0.05; ** p < 0.01; *** p < 0.001; **** p < 0.0001. P values for samples without significance are indicated. (b) Clustering larvae were removed, food color labeled and transplanted back and monitored. CS larvae spend about an hour in CS or P clusters. sgstai larvae spend about twice this time. (c) Larvae synchronization of CS into P or sgstai into P transplants. CS synchronize their movements by about 0.5 seconds, out of a 2-second locomotion cycle. This is about the same as CS into CS. sgstai synchronize less and are close to that of blind or naïve larvae. (d) Clusters in various strains and mixes were monitored for their average size. The number of larvae in each cluster is about 7 animals. Blind GMRhid larvae have reduced clustering but their clusters are about the same size. Only 3 sgstai clusters were seen and these have about the same size as other clusters. (e) Clusters in 3d were monitored for their average lifespan. All are about 3 hrs with the exception of 10sgstai30P and 10GMRhid30P. Blind GMRhid reduces host cluster lifespan 4 fold from 156’ to an average of 41’. sgstai raises the life span about 2.5 fold to 298’
Comments (0)