
Remember me
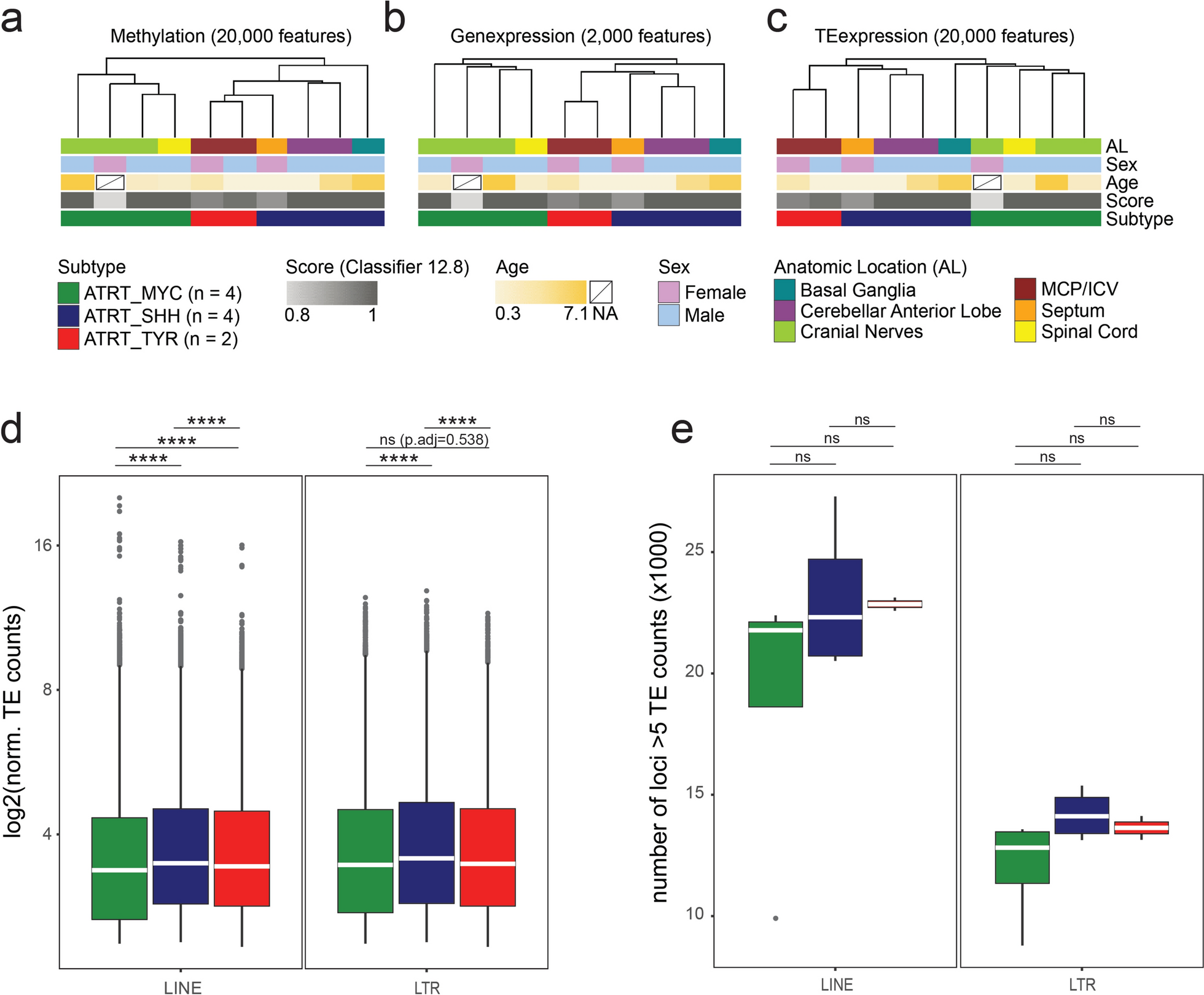




To screen the potentially effective drugs for PitNETs, we used a panel of 9 patient-derived PitNET primary cell cultures (Additional file 1: Table S1) for screening. Among these agents are different inhibitors for well-known oncogenic targets, such as Phosphoinositide-3-kinase (PI3K), mTOR, proteasome, HDACs, and mitogen activated protein kinase kinase (MEK), covering over 181 distinct mechanisms of actions (MoAs, Additional file 1: Table S3). Based on the reduction of cell viability, the screening generated 20,736 single-agent dose response signatures, resulting in a coherent database of PitNET vulnerabilities. Agents inhibiting at least 66.67% (6/9) patient-derived PitNET primary cell cultures by 50% were classified as ‘hits’. A total of 16 hits were identified (Fig. 1A, Additional file 1: Table S4). As shown in Fig. 1B, a total of 13 MoAs relevant to PitNET pathogenesis were enriched among the screened hits, including HDACs, PI3K, NF-κB, gp130, and others (Additional file 1: Table S5). Agents from these mechanistic classes demonstrated a relatively wide potency range, with HDACs inhibitors being the most potent drug class. Furthermore, we assessed the efficacy of agents targeting previously reported therapeutic MoAs to PitNETs [12], including HDAC, VEGF, RGF, FGF, PI3K, Akt, mTOR, Raf, MEK, ERK, Notch, Hedgehog, and CDK in MMQ, GH3, and ATt-20 cell lines using the HTS Library. As shown in Fig. 1C, Only HDAC was the therapeutic target for all three pituitary adenoma cell lines (p < 0.05).
Fig. 1
Identification of drug candidates through high-throughput drug screening in PitNETs. A Heat-map representation of drug activities for multiple PitNET cells screened through the HTS library. The gradual shift of color from green to red indicates the primary cell survival percentage from small to large. B Mechanistic drug classes enriched among the 16 ‘hits’ selected based on consistent potency across multiple PitNET primary cell cultures. C Left: An overview of growth inhibition of 3 pituitary adenoma cell lines by compounds targeting various MoAs. Right: A Venn diagram of the left results. D The cell survival of MMQ and GH3 cells for selected agents from the key enriched mechanistic classes HDACs
To verify the anti-tumor effects of HDACIs on PitNETs, the top 5 HDACIs, including Panobinostat, Quisinostat, Givinostat, Dacinostat, and Fimepinostat (CUDC-907), were used on GH3 and MMQ cells. Consistent with the HTS data, all these HDACIs strongly inhibited GH3 and MMQ cell proliferation at low concentrations, with IC50 values in the nanomolar ranges (Fig. 1D). Notably, Panobinostat displayed the lowest IC50 values for both MMQ (0.0615 μM) and GH3 (0.09575 μM) cell lines at 24 h, therefore it was chosen for further functional and mechanistic studies.
Panobinostat conferred anti-PitNET effects both in vivo and in vitroTo further verify the anti-tumor effects of Panobinostat in PitNETs, we first establish xenograft mouse model by implanting GH3 cells in the flank of nude mice. As shown in Fig. 2A, Panobinostat markedly inhibited the xenograft tumor sizes and weights (p < 0.05). In vivo TUNEL staining assays revealed that Panobinostat induced obvious cell apoptosis in xenograft tumors (Fig. 2B). Although it has been reported that Panobinostat has adverse side effects, mainly including diarrhea, pneumonia, and neurotoxicity [13]. Herein, we did not observe obvious mice diarrhea symptoms during the experiment. Moreover, Panobinostat did not affect the body weight, induced pneumonia or neurotoxicity of the mice significantly (all p > 0.05, Additional file 1: Figure S1A–E).
Fig. 2
Panobinostat demonstrated therapeutic efficacy in PitNETs. A Upper: Representative images of xenograft tumors from mice treated with control vehicle or Panobinostat. Down: Tumor volume growth curves and tumor weights in different treatment groups. n = 6; *p < 0.05, **p < 0.01. B Representative images of TUNEL staining from xenograft tumors. C The inhibitory responses of 18 primary PitNET cell proliferation by Panobinostat. D The cell survival of MMQ and GH3 cells after Panobinostat treatment at different dosage and time points. E The levels of PRL and GH secretions after Panobinostat treatment at different dosage at 24 h. n = 3; *p < 0.05, **p < 0.01. F Representation of colony formations in MMQ and GH3 cell lines. G Representation of Anneix staining in MMQ and GH3 cell lines
To test whether Panobinostat may inhibit human PitNET growth, we cultured the primary tumor cells from 18 PitNET patients, including PIT1-lineage (n = 7), TPIT-lineage (n = 1), SF1-lineage (n = 8), No distinct cell lineage (n = 2) (Additional file 1: Table S1). As shown in Fig. 2C, the growth of 13 of the 18 primary PitNET cultures was suppressed by 1 μM Panobinostat at 24 h, showing a total effective rate was 72.22%.
We next used two rat pituitary tumor cell lines, MMQ and GH3, to evaluate the anti-tumor effect of Panobinostat in vitro. The MTS assay showed that Panobinostat significantly suppressed MMQ and GH3 cell viability in a time course (Fig. 2D). The dose of 0.1 μM showed the most potent cytotoxicity at 24 h; while increase of dosages attenuated the cytotoxicity in both MMQ and GH3 cells. Also, Panobinostat markedly suppressed the PRL and GH hormone secretions from MMQ and GH3 cells (p < 0.05, Fig. 2E). Consistent with the cell viability results, Panobinostat significantly suppressed both MMQ and GH3 cell colony formations (Fig. 2F). As HTS evaluated growth inhibition rather than direct tumor cell death, the annexin V staining assays were used to measure apoptosis. Indeed, Panobinostat induced apoptosis in both MMQ and GH3 cells at 24 h (Fig. 2G). These data reveal that Panobinostat effectively inhibits pituitary adenoma cell growth and hormone secretion.
Panobinostat-treated PitNET cells exhibit altered transcriptional profilePrior studies suggested that Panobinostat was a nonselective pan-HDAC inhibitor targeting all four classes of HDACs; however, the exact mechanisms differ [14]. To explore the molecular mechanisms underlying Panobinostat on PitNETs, we performed transcriptome-wide RNA-sequencing analysis in GH3 and MMQ cell lines and 3 primary PitNET cell samples from patients (Additional file 1: Table S1) with or without 24 h treatment of 0.1 μM Panobinostat. We identified expression changes of 2299, 3895, and 1567 genes in GH3, MMQ, and primary cells respectively, upon Panobinostat treatment (Fig. 3A). To identify which pathways were affected by Panobinostat, we performed gene set enrichment analysis (GSEA). As shown in Fig. 3B, the Panobinostat-treated GH3, MMQ, and primary cells shared some same pathway patterns. Strikingly, the top 3 gene sets which were downregulated by Panobinostat were Nrf2-related signaling (all p < 0.05; Fig. 3B, C). In addition, Nrf2 downstream antioxidant genes, including NQO1, SLC7A11, GCLM, and SQSTM1, were downregulated as well in GH3, MMQ, and primary PitNET cells following Panobinostat treatment (Fig. 3D). To test whether Panobinostat can inhibit Nrf2 activation, we detected the changes in Nrf2 DNA binding activity in vitro. As shown in Additional file 1: Figure S2, Panobinostat inhibited Nrf2 DNA binding activity in a dose-dependent manner in both MMQ and GH3 cells (both p < 0.05). Interestingly, Panobinostat did not induce a significant inhibition in HDAC pathway, whose family members were highly expressed in different PitNET subtypes (Additional file 1: Figure S3). Specially, Panobinostat only slightly downregulated HDAC4 expression with no statistical significance (p > 0.05), and even slightly upregulated other HDACs (Additional file 1: Figure S4). These data suggest that Panobinostat may confer anti-tumor effects in PitNETs mainly via inhibiting Nrf2-mediated signaling, rather than directly affecting the expressions of HDACs.
Fig. 3
Transcriptome-wide RNA-sequencing assays to identify on-target ability of Panobinostat in PitNET primary, MMQ, and GH3 cells. A Transcriptome strategy of RNA sequencing conducted on different PitNET cells exposed to Panobinostat for 24 h is shown in schematic diagram. MMQ, GH3, and primary cell groups contained 3 replicates, respectively. B The common core-enriched signaling pathways changed following Panobinostat treatment. C Single GSEA was used to analyze the targeted signaling pathway. Normalized enrichment score (NES) indicated the analysis results across gene sets. D The gene expressions of Nrf2 and its downstream genes in PitNET cell lines following Panobinostat treatment by RNA sequencing
Nrf2 was highly expressed in human PitNETsTo confirm the potential involvement of Nrf2 in PitNETs, we analyzed the RNA-seq data on a total of 180 different PitNETs from our single expert center. Our data revealed that in general, the Nrf2 transcripts were highly expressed among almost PitNET subtypes (Fig. 4A). Also, the RT-PCR and Western blot analysis were used to detect Nrf2 mRNA and protein expressions in human normal pituitaries and PitNETs. As shown in Fig. 4B, C, Nrf2 mRNA and protein expressions were markedly higher in the PitNETs when compared to that in the normal pituitaries, respectively (p < 0.05). Furthermore, IF and IHC staining were employed to determine the relative changes in Nrf2 from the clinical PitNET specimens. As shown in Fig. 4D, E, few and abundant Nrf2-positive cells were observed in the normal pituitaries and PitNETs, respectively. To further explore the correlation between Nrf2 expression and PitNET clinical characteristics, we analyzed the Nrf2 expressions using IHC staining on 32 tumor samples from PitNET patients, whose clinical information were summarized in Additional file 1: Table S1. Notably, there was a significantly positive correlation between Nrf2 expression and tumor size, invasion, and recurrence (p < 0.05, Table 1).
Fig. 4
Nrf2 was highly expressed in PitNETs. A The gene expressions of Nrf2 in PitNETs by RNA sequencing. n = 180. B Validation of Nrf2 mRNA expressions in PitNET specimens. *p < 0.05. C Validation of Nrf2 protein expressions in PitNET specimens. D Representative IF images of Nrf2 staining in PitNET specimens. E Representative IHC images of Nrf2 staining in PitNET specimens
Table 1 Correlation of the expression of Nrf2 with clinicopathological features of 32 PitNET patientsNrf2 promotes PitNET growth and hormone secretionTo further investigate the effects of Nrf2 on PitNET development, we manipulated its expression in MMQ and GH cells. Over-expression (OE) and knockdown (KD) of Nrf2 were confirmed by western blotting. As shown in Additional file 1: Figure S5, si-R2 was selected in the following studies. The MTS assay showed that KD of Nrf2 significantly suppressed the proliferation of both the MMQ and GH3 cells, while OE of Nrf2 markedly promoted cell proliferation in a time-dependent manner (Fig. 5A, p < 0.05). ELISA assays showed that inhibition of Nrf2 expression significantly suppressed the PRL and GH levels in the culture medium from both the MMQ and GH3 cells. Conversely, OE group significantly increased PRL and GH secretion compared to the control group (Fig. 5B, p < 0.05). Consistent with the cell proliferation results, the colony formation assays also showed that KD and OE of Nrf2 markedly inhibited and promoted the numbers of MMQ and GH3 colony formations, respectively (Fig. 5C, both p < 0.05).
Fig. 5
The role of Nrf2 in the pathopoiesis of PitNETs. A The inhibitory responses of proliferation for different Nrf2 expressions in MMQ and GH3 cells. n = 3; *p < 0.05, **p < 0.01, ##p < 0.01. B The inhibitory responses of PRL and GH hormone secretions for different Nrf2 expressions in MMQ and GH3 cells. n = 3; *p < 0.05, **p < 0.01, #p < 0.05, ##p < 0.01. C Colony formation in MMQ and GH3 cell lines. n = 3; *p < 0.05, **p < 0.01. D Left: Representative images of xenograft tumors from mice with different treatment. Right: Tumor volume growth curves and tumor weights of nude mice in different treatment groups. n = 5; *p < 0.05, **p < 0.01, #p < 0.05, ##p < 0.01
Next, the above described Nrf2 OE, Nrf2 KD, and negative control (NC) GH3 cells was injected into nude mice to generate xenograft models. Similar to the results of the in vitro experiments, the mice received Nrf2 OE cells had bigger tumors, while the mice received Nrf2 KD cells had significantly smaller tumors comparing to the mice received the control cells, respectively (Fig. 5D, p < 0.05).
Nrf2 inhibition and Panobinostat demonstrate efficacy as combination treatment in PitNET growth in virto and in vivoGiven our RNA-seq results showing the potentially modulatory effect of Panobinostat on Nrf2 signaling, Nrf2 OE and KD GH3 and MMQ cells were employed. As shown in Fig. 6A, OE of Nrf2 significantly diminished Panobinostat-induced inhibition of MMQ and GH3 cell proliferation; while KD of Nrf2 markedly increased Panobinostat-induced inhibition of proliferation of MMQ and GH3 cells. Moreover, much lower levels of PRL and GH were detected in the culture media of MMQ and GH3 cells with Nrf2 KD comparing to those from the control group after Panobinostat treatment; but the levels of PRL and GH were significantly higher in the Nrf2 OE GH3 cells (p < 0.05, Fig. 6B). Similarly, KD of Nrf2 enhanced the suppression of colony formation and promotion of cell apoptosis in MMQ and GH3 cells after Panobinostat treatment. In contrast, OE of Nrf2 alleviated Panobinostat-mediated inhibition of colony formation and induction of cell apoptosis in MMQ and GH3 cells (Additional file 1: Figure S6A–C).
Fig. 6
The role of Nrf2 in the cytotoxicity mediated by Panobinostat. A The inhibitory responses of proliferation for different Nrf2 expressions with Panobinostat administration in MMQ and GH3 cells. n = 3; *p < 0.05, **p < 0.01. B The inhibitory responses of PRL and GH hormone secretions for different Nrf2 expressions with Panobinostat administration in MMQ and GH3 cells. n = 3; *p < 0.05, **p < 0.01. C Tumor volume growth curves and tumor weights of nude mice in different treatment groups. n = 5; *p < 0.05, **p < 0.01, ##p < 0.01. D Left: Representation of immuno-blots for different Nrf2 and NQO1 expressions in different groups. Right: Bar graph of densitometric analysis. n = 3; *p < 0.05, **p < 0.01, #p < 0.05, ##p < 0.01. E Redox changes evidenced by the levels of luminol CL, lucigenin CL, MDA content, and total SOD activity in each group. n = 3; *p < 0.05, **p < 0.01, ##p < 0.01. F Representative IHC images of H&E, 8-OHdG, Nrf2, NQO1, and caspase-3 staining in different treatment groups. G Representative images of TUNEL staining in mice PitNET specimens. H Representation of immuno-blots for different apoptosis-related protein expressions in different groups
Notably, there were no compounds targeting Nrf2 inhibition in our HTS libraries. To further test the anti-tumor effect of combination of Panobinostat with Nrf2 inhibition, ML385, a special Nrf2 inhibitor, was used. As Fig. 6C, ML385 significantly decreased the GH3 xenograft tumor sizes and weights compared with vehicle treatment (p < 0.05). Moreover, the tumor sizes and weights were much smaller in the Combination group compared to those from the Panobinostat group (p < 0.05). To further investigate the effects of Panobinostat, used alone or in combination with ML385 on redox homeostasis in vivo, we detected the protein expressions of Nrf2 and its downstream representative target antioxidant gene NQO1 by Western Blot analysis and the levels of oxidative stress by the CL assays of luminol and lucigenin, MDA content, and total SOD activity assays. As shown in Fig. 6D, E, the administration of ML385 enhanced Panobinostat-induced downregulation of Nrf2/NQO1 protein expressions and total SOD activities, and upregulation of luminal CL, lucigenin CL, and MDA content in the GH3 xenograft tumors (p < 0.05). IHC staining also showed that the 8-OHdG-positive cells were increased, and Nrf2- and NQO1-positive cells were decreased in the xenograft tumor samples after Panobinostat or ML385 administration; however, the combination group showed much more 8-OHdG-positive cells, and less Nrf2- and NQO1-positive cells compared to the Panobinostat or ML385 group, respectively (Fig. 6F). As for cell death, the number of TUNEL- and caspase-3-positive cells, the protein levels of PARP1, Bax, and caspase-3 were increased in the Combination group compared with those in the Panobinostat group. While the Bcl-2 protein expression was lower in the Combination group compared with that from the Panobinostat group (Fig. 6F–H). Together, the data suggest that the combination of Panobinostat and Nrf2 inhibitor ML385 has a synergistic effect, and effectively inhibits the MMQ and GH3 cell growth and hormone secretion, induces apoptosis, and enhances oxidative stress in virto and in vivo.
Previous data indicated that Akt-mediated phosphorylation of mTOR functions downstream of S6K1/4EBP1 to govern the activation of Nrf2 [15, 16]. Herein, Panobinostat decreased the phosphorylation of Akt, mTOR, and 4EBP1 in a dose-dependent manner in both MMQ and GH3 cells, without affecting S6K1 phosphorylation (Additional file 1: Figure S7). These data indicate that Panobinostat markedly inhibit Nrf2 activation, at least in part, via the Akt/mTOR1/4EBP1 phosphorylation.
Comments (0)