The aim of the current study was to utilize the amplitude of the mismatch negativity (MMN) component to assess auditory sensory memory for duration in individuals with CLN3 disease, on the premise that this easy-to-test neurophysiological marker might be sensitive to subtle changes in auditory cortical processing in this progressive neurodevelopmental disorder. Linear mixed effect analysis pointed to an MMN that was intact in the clinical group for medium presentation rates (900 ms SOA), reflecting a generally preserved ability to discriminate auditory duration deviance and to establish auditory sensory memories. However, the MMN was quite clearly compromised at longer (slower) presentation rates (i.e., the 1800 ms SOA) as greater demand was placed on the sensory memory system, in line with our main hypothesis. Results also suggested that at the most rapid stimulation rate (450 ms SOA), the MMN was weaker in the CLN3 cohort, an effect that was not predicted. Finally, we found that age significantly predicted neurophysiological correlates of sensory memory in CLN3 disease – that is, that the MMN showed a progressive reduction in amplitude with increasing age (i.e., disease progression), exactly opposite to what was observed in TD control participants.
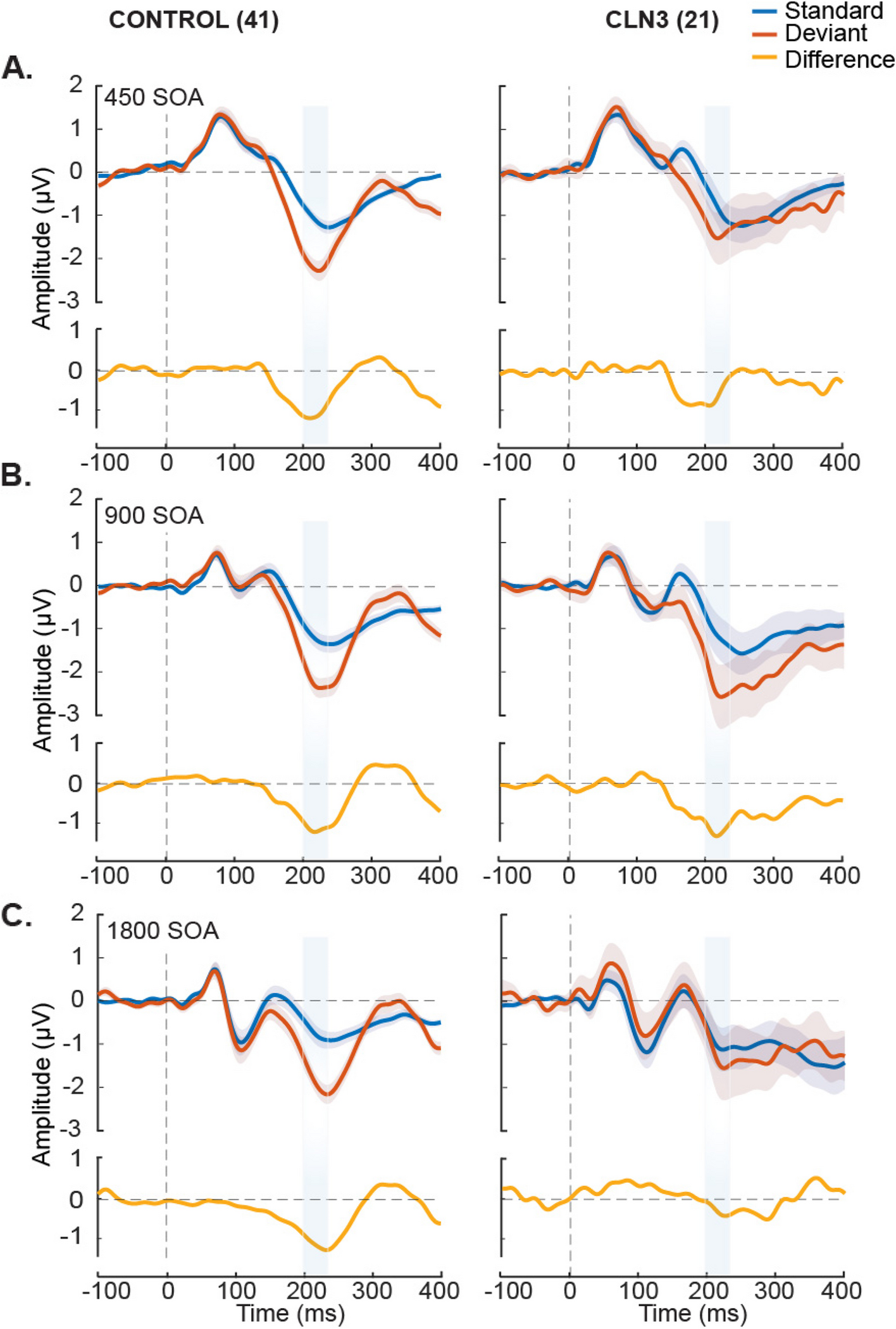
In what follows, we describe these results in more detail. First, there was a significant positive relationship between TD and CLN3 participants demonstrating that overall neurophysiological responsivity was relatively comparable across groups as a function of condition, although the absolute group MMN means were significantly different between each cohort. In addition, the a priori hypothesis that age would be significantly related to the MMN effect was supported as the overall fit of the model improved when age was added as a covariate. Additionally, there was a main effect for DEV vs STD trials which further demonstrated that there was an overall generalized MMN effect across both participant groups. A key finding was that presentation rate (i.e., the variation in SOA) was a significant predictor of participant group, indicative of an interaction between groups as a function of rate of presentation. When exploring this further using age as a fixed effect, the results showed that there was only a significant difference between the fast and medium SOAs (i.e., 450 ms and 900 ms), but not between the fast and slow SOAs (i.e., 450 ms and 1800 ms) or slow and medium SOAs (i.e., 1800 ms and 900 ms). This suggested that the MMN in CLN3 participants was equally disrupted in the fast and slow presentation rates and the largest gap in auditory sensory memory performance was between the fast and the medium presentation rates.
Next, using a within-subjects model to explicitly test for the MMN effect in the CLN3 group, no significant difference between DEV vs STD trials (i.e., MMN effect) was observed at both the fast and slow presentation rates, whereas there was a robust MMN effect at the medium SOA rate. In partial contrast to the LME results, exploratory post-hoc cluster statistics did show a significant MMN effect at the fast presentation rate in CLN3, whereas both statistical approaches showed no evidence of an MMN at the slowest rate. This difference in absence and presence of MMN effect at the fastest presentation rate (SOA = 450 ms) is most likely due to the inherent methodological differences in these statistical approaches [66]. The computation of cluster-based statistics takes into consideration adjacent temporal and spatial information and uses cluster-based correction methods to account for multiple comparisons, while the LME approach relies on specific time-windows of interest at a fixed region of interest (i.e., pre-specified electrodes). Additionally, the LME model used age as a covariate. Lastly, it is worth noting that while the MMN was not strictly significant at the 450 ms SOA in CLN3, it did approach conventional levels of significance in the LME (p = 0.053) and Bayesian analysis pointed to moderate evidence for an MMN at this presentation rate. When focusing on the TD cohort, the MMN was clearly present and highly stable across all SOAs, as expected.
Prior work has shown that the strength of the MMN is highly dependent on stimulation rate, with reduced MMN responses observed at slower rates [41, 42, 67]. Current understanding of this phenomenon is that the strength of the auditory sensory memory depends on a temporal integration window, such that establishment of a robust sensory memory depends on the presentation of a number of standards within this window, against which the deviant will ultimately be compared. Perceptually, this is very obvious in a design such as the one used here. At rapid rates of presentation (e.g., (SOA of 450 ms)), the duration deviant pops out strongly from the rapid stream of standards, whereas when the rate of presentation is slowed (SOA 1800 ms here), this pop-out is diminished. In extremis, the reader can readily imagine that if the standard tones were presented once per minute or at even longer lags, it would become very difficult to determine a duration deviant relative to these sporadic standards, and this would certainly not be achieved automatically (pre-attentively). The fact that duration MMN is absent at the slowest and most demanding presentation rate here in CLN3 disease, may point to the early stages of a breakdown in automatic detection and integration of these stimuli in auditory sensory memory. It is also worth pointing out here that the presence or absence of an MMN during passive tasks is known to correspond closely with behavioral performance when individuals are asked to actively discriminate the deviants in follow-up behavioral studies. Only deviants that can be discriminated above chance levels are found to also evoke MMN responses [67, 68].
We did not behaviorally assess auditory discrimination abilities of the participants given the associated loss of vision, speech, and motor decline. Many of the participants with CLN3 disease would not have been able to perform the task. Rather, we employed the passive MMN design to assess the evoked neural activity. It will fall to future work to determine what the perceptual and cognitive implications of this breakdown are [10, 11, 69]. However, prior work has shown that weakened ability to sustain information in sensory memory can reflect cognitive deterioration in various clinical conditions [42, 45, 46]. It will also be of significant interest to further investigate the duration-evoked MMN at even slower presentation rates. This may better reveal the extent of this difference, and it remains to be determined whether this difference is peculiar to the feature of duration or if it will also be evident for other basic auditory features such as pitch, loudness and location. Manipulations of presentation rate are not the only way in which the auditory sensory memory system can be parametrically manipulated. Whereas the presentation rate manipulation used here is presumed to test the temporal integration window of the MMN system, the sensitivity of the system can also be assessed by manipulating the extent to which the deviant stimulus differs from the standards. Here, a deviant of 180ms was used against a standard tone of 100ms, which represents a large and highly discriminable duration change known to evoke large amplitude MMN responses in neurotypical controls [40]. By parametrically manipulating the extent of the duration deviance, prior work has shown that the amplitude of the MMN tracks with the size of the difference, such that at small differences (e.g. 130ms versus 100ms), the MMN is highly diminished or even absent in neurotypical controls [40].
There are parallels between the current findings of diminished MMN responses at slower presentation rates and prior work in other rare neurodevelopmental diseases, specifically Rett Syndrome (RTT) and Cystinosis [12, 13]. In Rett participants, for example, the duration-evoked MMN was only detected when stimuli were presented at the most rapid presentation rate of 450 ms SOA, and unlike the CLN3 disease participants reported here, no MMN was evident at the 900 ms SOA, nor at the 1800 ms SOA, suggesting a more severe disease course in this population. Likewise, participants diagnosed with Cystinosis, another of the rare lysosomal storage disorders, produced robust MMNs comparable to those seen in TDs only in response to the fastest presentation rate (i.e., at 450 ms SOA) [13], with clear atypicalities in the MMN at the two slower rates 900 ms SOA and 1800 ms SOA). Taken together, these data suggest that the duration-evoked MMN may be a sensitive measure of disease severity across a number of neurodevelopmental disorders.
An unanticipated finding here was the weakened MMN response in CLN3 disease at the fastest presentation rate (i.e., the 450 ms SOA). This is the rate at which one expects the most robust MMN to be produced, whereas it was at the medium rate (900 ms SOA) that this occurred in CLN3. Since this was not explicitly predicted, the effect warrants replication in an independent cohort before any strong conclusions can be drawn. Nonetheless, these data suggest that there may be an emerging deficit in the ability to generate auditory sensory memories for duration at rapid presentation rates in CLN3 disease.
Clinically, some of the most striking differences observed in individuals with CLN3 disease are in memory, attention and speech functions [5, 6, 10, 11]. This cognitive decline generally begins around the time of onset of vision impairment, but continues to progress over years, even after vision loss is maximal [9, 70]. To date, quantitative characterization of these differences has not been well-defined [11]. As such, the relationships between cognitive impairments and other clinical features of CLN3 disease are not yet well understood. For instance, the onset of visual decline and of cognitive deterioration have been a subject of debate [6]. It is generally accepted that the onset of observable cognitive decline begins within two years of the onset of visual decline [6, 10]. This has been shown in some individuals with CLN3 disease, while in others, this decline seemed to precede visual deterioration or even emerge at a much later stage [71,72,73,74]. These inconsistencies in the manifestation of the onset of cognitive decline were taken to highlight the importance of careful acquisition of patient history in those suspected to have CLN3 disease [6]. Similarly, understanding the extent of cognitive regression in CLN3 disease is an important component in identifying reliable neurophysiological biomarkers of this disease. As far as we know, the use of electrophysiological assays to evaluate cognitive abilities including attention and memory has not yet been leveraged in this population. The current work serves as a good first step in exploring and developing objective neural markers of pathology (biomarkers) that can be easily carried out noninvasively throughout the progressive stages of CLN3 disease.
Genetically manipulated mouse models of disease are remarkably powerful research tools, providing essential insights into the neurobiological substrates of neurodevelopmental disorders like CLN3 disease [75,76,77], and yet many of the outcome measures used to quantify or track disease progression in a mouse cannot be meaningfully applied in humans. Obviously enough, invasive electrophysiological recordings, ubiquitous in model systems work, are not feasible in humans. Similarly, many of the behavioral assays used to assess disease progression and severity in a mouse are only loosely related to human behaviors [78], and higher-order functions such as cognitive control and language cannot be readily interrogated. Establishing objective neurophysiological markers of disease progression in human patients is a crucial step towards bridging this inter-species translational divide. In humans, measures of brain electrophysiology are almost exclusively made using non-invasive scalp recordings that assay the activity of large distributed neuronal ensembles across the entire brain (i.e., circuit-level analysis). In mouse models, typical assays involve single or multi-unit neuronal recordings in vivo (usually in anesthetized preparations) or in vitro slice preparations where synaptic plasticity can be assessed. Again, while the approaches used in each species are certainly powerful in their own right, the researcher is mostly left to infer or speculate about correspondences across species. However, ERP markers like the MMN can be readily recorded in mice using wholly similar, if not identical experimental procedures [79, 80]. It will be important to determine going forward whether in mouse models of Batten disease, the MMN phenotype seen here can be recapitulated. If so, it will present as an excellent cross-species neuromarker.
Study limitations
A few limitations of the current study need to be acknowledged. Given that auditory responses continue to mature with typical development [43, 81], the relatively wide participant age-range is a limitation and follow up studies will ideally work within more delimited age-ranges. Of course, given that CLN3 disease is a rare disease, recruitment within restricted age bands is very challenging. In addition, although age was correlated with MMN amplitude in the TDs, it was not associated with manipulations of stimulus rate. This suggests that the differences seen among groups as a function of presentation rate were not affected by age, but rather, represent frank differences in brain function in CLN3 disease. It will be crucial for future work to follow up with parametric studies to assess the limits of the auditory sensory memory system in CLN3 disease for these and other fundamental auditory features (i.e., frequency, duration, location, and loudness) and their implication for higher-order cognitive processing. Future studies should also follow up with the evaluation of the relationship between ERP measures and the four disease stages based on the CLN3SS, with more patients representing each disease stage. In this study, exploring the effects of CLN3 disease stage on MMN amplitudes as a function of SOA while controlling for age proved to be the best model for LME analysis. Although including age as a covariate in the LME model improved performance, the outcomes should be interpreted with caution due to the relatively restricted sample size in each of the stages of CLN3 disease. Again, recruitment within restricted disease stages just as with restricted age bands is very challenging given that CLN3 is a rare disease. We did not include biological sex as a variable in our analyses, and this may be of importance in future work given the reported sex differences in symptom severity and progression in CLN3 disease [82,83,84]. The current study was not adequately powered to examine this variable (9 females versus 12 males in our CLN3 cohort). It is worth pointing out though that there is no clear evidence for biological sex differences in the generation of the MMN [85]. Lastly, non-invasive recordings such as those conducted here are limited in their ability to shed light on the mechanisms by which CLN3 protein dysfunction leads to auditory cortical processing differences. Work using similar paradigms in murine models of CLN3 disease will be highly instructive in this regard [86, 87, 77].









Comments (0)